植物プランクトンの生産性に影響を与える主な要因。 海洋の植物プランクトンはますます減少しています
「ロシア科学アカデミー」
UDC 574.583(28):o81 +574.55:58.035
ピリンル インナ ロプショヴナ
内陸水域における植物プランクトンの生産性の要因として
03.00.16 - エコロジー
科学レポートの形式で生物科学の学位を取得した科学者を獲得するための競争。
サンクトペテルブルク 1995
この研究は、その名にちなんで名付けられた内水生物学研究所で実施されました。 II. D. パパシュシャ RAS。
公式対戦相手:
生物科学博士 Lavrentyeva G.M. 生物科学博士 Bulyon V. V. 生物科学博士 Raspopov I.M.
主導機関: ヴォルガ流域生態学研究所 RAS "....
防衛は「×」で行われます。 /「1995 年 4 月」時。 ロシア科学アカデミーの湖沼科学研究所(198199、サンクトペテルブルク、セヴァスタノヴァ通り、9)での専門評議会D 200.10.01の会議にて。
この論文は科学報告書の形式で、ロシア科学アカデミー湖沼科学研究所の図書館に所蔵されている。
専門評議会の科学書記
生物科学の候補者
M.A.ベロバ
導入
研究の関連性
水生生態系の状態の悪化に関連して、天然水の水質の形成、特に貯水池に一次有機物と酸素を補充するプランクトンの光合成に影響を与えるプロセスの研究が行われています。ますます重要になっています。 水生環境で起こるこのプロセスにとって最も重要な要素は光です。 これは、太陽光のプランクトンへのアクセスが限られているためであり、その大部分は水やneiiに含まれる物質によって吸収および散乱され、光感作細胞に到達しません。 さらに、透過放射線のスペクトル組成は水中で変化します。主に光合成に使用される赤色と青色の光線は最も強く保持され、緑色の光線はより完全に透過されます。 その結果、プランクトン性フィトセノーシスは、光合成に必要な炭酸塩と水の化合物、ミネラル栄養成分、およびかなり安定した周囲温度の点で陸生のものよりも有利であるが、エネルギー源の面では不利な立場にある。 そして、空気によって弱められた太陽光を比較的ほとんど受けない地球の表面では、植物が光エネルギーに不足しておらず、その光合成が他の要因によって制限されている場合、水中でのこのプロセスは光によって最も制限されます。
陸生フィトセノーシスの研究者によって開発された植生の光合成生産性の理論では、作物形成の要因として太陽放射エネルギーが非常に重要視されています (Nichiporovich 1956, 1908)。 水生生物学者が水生生態系の一次生産物を評価する際、特に淡水域を研究する場合にはこの要素に注目することは比較的まれですが、水光学の観点から見ると多様性が大きいため、この種の研究が最も必要とされています。 したがって) 水中光合成の光因子に関する特別な研究は、30 年代に湖で始まり (Schomer、J934; Schomer、Juday、935: Manning、Juday、1941、)、50 ~ 80 年代に取り上げられました (Talling、¡ 957、1971、1982;Tilzer、Schwarz、1976;Tilzer、984;Ganf、1975;Jewson、1976、1977;Kirk、1977、1979 - 引用;Kirk、1983;Roemer、Hoagland、1979 引用:カーク、 1983; Megard et a! さらに、それを、水域の富栄養化の問題により、最近の陸水学的研究が逸脱した植物プランクトン生産性の生物起源の研究と比較すると、
水中光合成の要素としての光の研究は、光エネルギーを捕捉する植物プランクトンの色素の研究と密接に関連しています。 主なものはクロロフィル「a」で、光合成を行うすべての植物と地域に特有の物質です。
ユニークなスペクトル特性を与えることで、残りのプランクトンから藻類を分離することなく、藻類のバイオマスを定量化することが可能になります。 光合成のプロセスに直接関与すると同時に、植物プランクトンの同化活動の指標としても機能します。 他の色素は、その多くが藻類に特有のものであり、深さが変化する光の条件下での水中光合成の生態を理解するために重要です。
光合成プランクトンの生物量を決定する際にクロロフィルを使用するというアイデア (Vinberg、1954、1960) は非常に有益であることが判明し、さまざまな種類の水におけるこの色素の研究に関する広範な研究の発展の基礎となった。連続モードで水中で直接分析したり、遠隔から分析したりする特別な分析方法を開発して、遺体を分析します。 クロロフィルの定義は、ほとんどの水文学研究に含まれており、クロロフィルは天然水の生産性と水質の指標と考えられており、環境上の「水域の監視」に不可欠な部分となっています。この色素への関心が急増しています。現代の水生物学において、クロロフィルが植物の光合成における役割を発見したのは少なくとも約 100 年前であることが判明しましたが、プランクトンによる太陽エネルギーの吸収を確実にするクロロフィルの特定の特性や、特に淡水域では、水中光の下での光合成における他の色素はほとんど存在しない(Tilzer, 1983; Ganf, et al., 1991)。
一方、浮遊藻類の光合成と太陽放射およびクロロフィルのエネルギーとの関係を特徴付けるパラメータは、一次植物プランクトン生産の決定および数学的モデリングのための広範な計算手法の基礎となっています。 特定の自然環境に最も適したこれらのパラメーターの値を知ることが重要です。 海洋学者は、自然実験に基づいた研究も含めて (Platt et al., 1980, 1990; その他)、この方向で非常に綿密な研究を行っています (Koblenz-Mishke, 1980; Koblenz-Mishke et al., 1985; 1987)。 淡水の水域では、そのような研究はあまり開発されておらず、水中光合成のモデル化に必要なパラメーターは主に理論的に、または文献情報源から見つかります (StraSkraba、Gnauck、1985)。
研究の目的と目的
主な連鎖は、淡水生態系における光合成および一次生産物の形成中の水中の光条件と植物プランクトン色素の含有量との関係を特定することで構成されていました。
この目的のために、以下の具体的な課題が設定された。 (1) 溶液の影響を考慮して、光学的に異なる淡水域における一般スペクトルおよび光合成活性領域の太陽放射エネルギーの侵入および浸透のパターンを研究する。
ゆるやかな色の物質、一般的な懸濁液および植物プランクトン細胞。 (2) 光合成バイオマスの指標としてのクロロフィルの含有量、他の植物プランクトン色素との比率を決定し、それらの空間的、季節的および経年的変化のパターンを研究し、研究された水域の栄養状態に関連する差異を評価する。 (3) 光条件とクロロフィルの量と比較して、植物プランクトンの一次生産レベルと太陽エネルギーの利用効率を決定します。 (4) 植物プランクトンの生産性と貯留層の富栄養化における年々変動における、地球に流入する太陽エネルギーの役割を評価する。 (5) 自然環境における多様な色素をもつ植物プランクトンの光合成の光依存性を研究し、それらを一次生産のモデル化に使用する可能性を研究する。
保護条項
I. 植物プランクトンとそれを捕捉するクロロフィルが利用できる光エネルギーの量によって、水域の一次生産レベルが決まります。 2. プランクトンによる植物生成の比較的単純な構造により、光合成の過程で強化された太陽光が貯留層に浸透するパターンが数学的に非常に簡単に記述されます。 3. これにより、機器測定された特性に基づいて植物プランクトンの一次生産量を評価するための計算方法の開発の可能性が開かれます。
科学研究
1. オリジナルの装置を使用して、淡水貯水池(ヴォルガ貯水池、プレシュチェゾ湖、オネガ湖)の広範囲で水中光合成活性放射線のエネルギーの最初の測定が行われました。 光学的に異なるタイプの水域への放射の浸透パターンが、氷下の期間を含む緯度と季節の側面で研究され、全太陽スペクトルにおける割合が評価されました。古典的なブーゲー法則で説明される、自然水域におけるスペクトル的に複雑な太陽光線束の特徴が明らかになり、数学的な方法でこの偏差を補償するパラメーターが見つかりました。緑化における他の浮遊物質に対する植物プランクトンの寄与透過放射線が測定されました。
2. ヴォルガ盆地、ラドガ湖とオネガ湖、ツンドラのハルベイ湖、川の貯水池と湖で植物プランクトン色素の含有量が初めて測定されました。 エニセイは、その誘導体の量におけるch.chozophylla "a"の割合と他のクロロフィルとの割合、および緑と黄色の色素の総量の割合を評価しました。」それらの空間分布のパターン、季節および長期変動が研究され、特定のデータに関する最初のデータが得られました。
4. ルイビンスク貯水池での研究中に、クロロフィル含有量に関する世界最長の長期観察シリーズの 1 つ (27 年間) が得られました。 その年次変動と異なる年の総括的な特徴との間の関係が示されています。 色素濃度のレベルが上昇する傾向が確認されており、これは貯留層の富栄養化と、この過程で地球に入る太陽放射のエネルギーの役割を示しています。
5. 多くの貯水池(規制前のV.オルガの特定地域、イワンコフスコエ貯水池、北ドヴィナ湖とハルベイ湖、プレシチェエヴォ湖、ラドガ湖)が、植物プランクトンの一次生産に関連して初めて研究された。
6. 研究された貯留層において、植物プランクトンによる太陽放射エネルギーの利用効率と、光条件およびクロロフィル含有量との関係が初めて評価された。
7. 自然環境における独自の実験に基づいて、生態学的および分類学的に多様な植物プランクトンの光合成の光依存性に関する新しいデータが得られました。 この依存性の分析的表現が与えられます。
8. 植物プランクトンの一次生産の指標の研究に応用できるいくつかの新しい方法論の開発が行われた: (1) 水中の光合成活性放射線を測定する装置が設計され、それを放射照度の単位で校正するための簡単な方法が提案された。光量測定法に基づく。 (2) 全抽出物中の植物プランクトン色素の分光測光分析が、国内の水生生物学研究の実践に導入された。 (3) 入力強度に基づいて植物プランクトンの一次生産量を決定するための計算方法が開発されました。 貯留層の日射量と水中光合成の光依存性。 (4) 貯水池および季節期間の平均植物プランクトン特性を計算する手順が改善されました。 (5) 正確な分野で一般的な間接誤差を評価する方法を使用して、測定誤差を計算し、植物プランクトンの生産性指標を平均するための独自のアルゴリズムが開発されています。
実用的な価値
植物プランクトンの光合成活動は水域の有機物と酸素の富化に関連しているため、この研究の枠組みの中で行われた研究は、自然の状態での天然水の品質を評価することを目的とした多くのプロジェクトの一部であり、さまざまな人為的影響下にあります。 これは、(1) ルイビンスクとイワンコフスキーの水質形成における浅海の役割を特定するために政府科学技術委員会 (GKNT) によって提案された研究です。
コナコヴォ州地区発電所の試運転後を含む、飲用イワンコヴォ貯水池の生態学的状態の評価による貯水池(1971~1973年、1973年の報告書)と貯水池ヴォルガ川 - ルイビンスク貯水池を含むバルト海系について、河川流量の再配分の問題に関連して(1976年から1985年、1980年と1985年の報告書)。 (2) 植物プランクトンの色素に基づいて貯水池の生産性を監視するための遠隔方法を開発することを目的としたルイビンスク貯水池での航空宇宙実験(1986~1990年、1990年報告)。 (3) 湖沼の環境モニタリング等の地方行政機関及び実務団体の指示に基づいて行う業務。 プレシチェフ (1986 ~ 1992 年、1986 年、1990 年、1992 年に報告)。 クイビシェフ貯水池の生物相に対する地面からの砂と砂利の混合物の除去作業の影響の評価(1990年から1991年、1991年の報告書)。 (4) 水域の環境モニタリングの実践においてこの方法を普及させることを目的として、水域中のクロロフィルの研究に関するクラスノヤルスク州立大学との創造的協力に関する協定に基づいて活動する(1986~1987年、1987年報告書)。 ) 環境プログラム「マンとブヌーファー」(プロジェクト No. 5) の枠組みの中で活動し、貯水池の一次生産レベルに影響を与える要因を特定する (1981 年から 1990 年の年次報告書、および 1986 年、1988 年、1991 年の統合報告書) )。
作品の承認
この研究の結果と主な規定は、貯水池の科学的生産に関する最初の会議で提示された(ミンスク、1960年:ソ連の他の貯水池における緑藻の役割の問題に関する全連合会議で(コロク、1960年)) *: 1. P, Sh. VI オーブとエネルギーと湖の貯水池に関する全連合陸水学会議 (Lisgvenichnoye-on-Bankale, |"64, 196"-). 1973, 1985); I (モスクワ、!%) 5] 、I (Kshshshev、197I.). V !ol! from i. 1986) a.ezlnkh VGBO: at 1 (Tolyatti. 1968) および II (Kor. ¡974 ¡.; ヴォルガ貯水池に関する会議と研究盆地; 貯水池の共同統合利用について (キエフ、1997); 火力発電所の下で加熱された貯水池の水文学と生物学に関する神経シンポジウムで (Eorok、197!): GTA TU (キエフ、1972)。) および XI! (Lnstvenichnoye-ia-Bankale、1984) 光線測定に関する全組合会議; エトロフィナス水域の問題に関する II 全組合シンポジウム (Zvesh!gorod、1977); II 全組合会議「生態学の問題」バイカル地方の」(イルクーツク、1982年)。 ヴォログダ地域における生物生産性、合理的利用、水域の保護の問題に関する会議にて(ヴォログダ、1978年)。 全連合科学会議「ソ連の大湖の天然資源」(レニングラード、1982年)にて。 内部資源の保護と合理的利用の問題に関する地域会議で
ロシア平原の中央部と北部の地域水域(ヤロスラヴリ、1984年)。 内陸水域の航空宇宙探査問題に関するIおよびI国際会議(レニングラード、1987年、1988年)において。 全連合学校セミナー「水生物学における定量的方法」(Borok、1988)にて。 イ・ヴェレシチャーギン・バイカル国際会議(リストヴェニチノエ・オン・バイカル、1989年)にて。 内陸水域におけるプランクトンの一次生産の研究(Borok、1989)および植物プランクトンの生産性の評価(Irkutsk、1992)に関する方法論会議で。 英国淡水生物協会の会議にて(ウィンダミア、1990年)。 ロシア艦隊創設300周年を記念した全ロシア科学会議にて(ペレスラヴリ・ザレスキー、1992年)。 大河川研究における環境問題に関する会議で(Tolyatti、1993)。 内陸水域における長期の水生生物観察に関する会議にて(サンクトペテルブルク、1994年)。 また、ロシア科学アカデミーの内水生物学研究所の勤務先でのセミナーや学術会議でも同様です。
出版物
研究結果は、10 冊の単行本と 65 冊の雑誌およびその他の記事の 15 セクションで発表されています。 彼の博士論文を擁護する前に、このトピックに関して 10 件の記事が掲載されました。
I. 資料と研究対象
この研究は、ヴォルガ貯水池とツンドラのハルベイ湖にある隣接する貯水池における植物プランクトンの光条件、色素含有量、光合成強度の研究結果に基づいており、生育期全体にわたって実施された(イワンコフスコエ貯水池 - 1958年、1970年 -) 1971、1973 - 1974; ルイビンスク貯水池 - 1958、1969 ~ 1973 年; クイビシェフ貯水池 - 1958; ベロエ湖 1976 ~ 1977; プレシチェエヴォ湖 - 1983 ~ 1985 年; ハルベイ湖群 1969)または特定の季節期間のルート調査(ヴォルガ川 - 195) 7 、1960; ラドガ湖とオネガ湖を含むヴォルガ・バルト海および北ドヴィナ水路の貯水池 - 1973; シェクスナ貯水池と上ヴォルガ貯水池 - 1979)。 イワンコフスキー保護区における植物プランクトン色素の観察は、1977 年から 1978 年にかけて湖で継続されました。 プレシチェエヴォ - 1991年まで、ルイビンスク貯水池では継続的な長期研究に移行し、それは現在も続いています。 植物プランクトンの色素の詳細な研究は、オネガ湖の国立公園 (1967 年から 1968 年) と川で実施されました。 エニセイ (1984-1985)。 ルイビンスク貯水池では、水域の遠隔光学センシング技術を開発するための航空宇宙実験の一環として、顔料含有量の小規模な変化が研究されました。
水域と時間(1986-1988)。 ルイビンスク貯水池 (1971 ~ 1972 年) とイワンコフスキー貯水池 (1973 ~ 1974 年) では、貯水池全体の一次有機物の濃縮におけるその役割を評価するために、浅いゾーンで一連の作業が行われました。 ルイビンスク貯水池と湖。 プレシチェエヴォは、氷河下時代の明るい環境の条件下で、植物プランクトンの発達に関する冬季観察を実施した。 ヴォルガ湖とシェクスニンスキー貯水池(i960、1979年)、ルイビンスク貯水池(1970年から1971年、1987年)、オネガ湖(1968年)とプレシチェエヴォ湖(1983年から1984年)へのルート遠征中に、太陽放射の研究に関する特別な作業が行われました。水への浸透と水中光合成の光依存性。 光合成活動はイワンコヴォ貯水池で研究されました。 クイビシェフ貯水池の火力発電所からの廃水の影響下で生成された、高温条件下(1970年から1971年)のプランクトン - 浚渫やその他の作業中に貯水池に入る鉱物懸濁液の植物プランクトンへの影響。 土壌除去に関するもの(1990~1991年)。
一部の資料は内水生物学研究所の職員(J1.B. Morokhovets、O.I. Feoktistova、N.P. Mokeeva、A.L. Ilyinsky、V.A. Elizarova、E.I. Naumova、V.G. Devyatkin、L.E. Sigareva、E.L. Bashkatovon、N.M. Mnneeva、L.G. )と共同で入手しました。 Korneva、V.L。Sklyarenko、A.N。Dzyuban、例えばDobrynin、M.M。スメタニンなど)およびその他の科学機関(V.A. Rutkovskaya、I.I。Nikolaev、M.V。Getsen、T.I。Letanskaya、I.S。関連する出版物この作業には、必要なデータを含む独自の記事が書かれているか、または独立した記事が使用されています。 T.P.は長年にわたり素材の収集と加工に常に参加してきました。 Zaiknna は、内陸水生物学研究所の藻類研究室の上級研究員助手です。 彼ら全員、コンピュータセンターと実験ワークショップのスタッフ、内陸水域生物学研究所の技術スタッフ、モエック大学、フスキー大学、サンクトペテルブルク大学、ニジニノヴゴロド大学、ヤロスラヴリ大学、ペルミ大学、カザフスタンの大学の学生たち、著者は、同研究所でインターンシップを行った方々に対し、研究の実施にご協力いただき、深く感謝の意を表します。
II. 研究手法
ライトモードの特性を検討する
植物プランクトンの主な生産における要因としての光体制の条件の研究は、通過する太陽の放射エネルギーが量的にだけでなく質的にも変化するという事実により、重大な方法論的困難を伴います。 スペクトル構成と角度
透過放射線の特性により、水中に入る放射線の強度は、地平線からの太陽の高さや雲の状態に応じて連続的に変化します。 理想的には、深度変化と光合成活性放射線の両方を時間の経過とともに合計した放射照度の単位で表される PAR (k = 380 ~ 710 nm) を記録できるデバイスが必要です。そのようなデバイスは存在しなかったためです (レポート... 、1965、1974)。 このデバイスは、半球からの放射、すなわち放射照度(コサインの法則に従う)を捕捉する一連のセンサーで構成されており、そのうちの 1 つは PAR (380 ~ 800 nm) をカバーする波長範囲で感度が高く、他のセンサーはこのより狭いセクションにあります。スペクトル領域 (480-800、600-800、680-800 nm)。 センサーのスペクトル感度は、真空フォトセル TsV-3 と光フィルター SZS-14 + BS-8 を組み合わせることで実現されます。光フィルターは 380 ~ 800 nm の全波長範囲をカバーし、SZS-14 + ZhS-17、SZS-14 + KS-10、SZS-14 + KS-19 - 残りの部分はそれぞれ徐々に先細になっています。 記録装置は、時間の経過に伴う放射線エネルギーを自動的に合計します。
放射照度の単位での装置の校正は、アクチノメーター法 (Berezkin、1932) に基づいて開発され、適切な光フィルターを備えたアクチノメーターによって測定される直接太陽放射に従って実行されます。 直接光線は、センサーに取り付けられたチューブを使用して、センサーに到達する全放射線の流れから分離されます。このチューブは、設計がアクチノメーターチューブと同様です (Pyrina、1965、1993)。
標準的な光量測定装置との比較を含め、水中フォトインテグレータを使用した長年の経験により、積分 PAR フラックスとこのスペクトル領域の狭いセクションの両方の測定結果が十分に高い精度で、フィールド条件での動作の信頼性が証明されています。 。 約30年前に作られた最初のサンプルは今でも機能しています。 さらに、Li-Cor や QSP (米国)、QSM (スウェーデン) などのフェーズド アレイの水中測定用のシリアル デバイスは現在存在しており (Jewson et al., 1984)、我が国の単一モデル (Semenchenko et al., 1984) も同様です。 、1971年;チェコ共和国、1987年)、まだアクセスできません。
植物プランクトンの一次生産量を決定するためのほとんどすべての実験は、貯留層に入る PAR スペクトルにわたって積分されたエネルギーの測定によって測定されました;この目的のために、フォトインテグレーター センサーが船の上部構造の上部または海に開いた高台に設置されました。海岸にある太陽、露光時間中の放射線の流入を記録。
貯水池内の光の状態を研究する場合、スペクトル感度が異なる水中フォトインテグレーターセンサーのセット全体を使用して、PARスペクトルのいくつかの部分で測定が実行されました。 センサーはフロートに吊るされた貯水池に発射され、フロートとともに船から10〜15メートル離れたところに移動されましたが、太陽の方向に細長い延長部分を備えたウインチを使用することはあまりありませんでした。 これらの測定の一部には、一般スペクトルの太陽放射の透過の観測が伴っていました。これらの観測は、1960 年に V.A. ルトコフスカヤ (1962、1965) によって、1979 年に T.N. によって行われました。 ポカチロワ (1984, ¡993)。 Onn は Yu.D. 日射計を使用しました。 Yashpievsky (1957) は水中測定に適しており、取り外し可能なウインチを備えた貯水池に浸漬されていました。 透過放射線のすべての測定は、機器の閾値感度により信頼性の高い測定値が得られるようになるまで、10 ~ 12 水平線の深さで実行されました。
植物プランクトンの光依存性に関する実験を行うために太陽放射の測定が使用される場合、試験サンプルの入ったフラスコが装置に取り付けられ、そのような装置は暴露中ずっと貯水池内に暴露されました。 これらの設備には、水の透明度に応じて計画された、光合成を決定するための複数のホライズンが装備されていました。 このようにして、サンプルに入る放射線エネルギーを完全に記録し、天然プランクトンの光合成の光依存性に最も適切なデータを取得することができました。
必要に応じて、水中の光の状態に関するデータを入手してください
多くの観測所では、時刻 t での到達に基づいて透過放射線を決定する計算方法に頼っていました。 透明牛 提案された F.E.、Api c D.I. を使用して Sekhki を焼きます。 Tolstyakov (1969) の公式、またはその後の洗練されたパリチット (Pyrina、1989)。
一般スペクトルの入射放射線のエネルギーからの izanmotrans.chod:: (l. = 380-710 im)、またはフォトインテグレーターによって記録された (l = 3íW-300 i, i)、およびその逆は、経験的係数 (Pyrnna) を使用して実行されました。 、1985)、pavnenmoeshまたは性格に関する関係、および晴れた天候で-太陽の高さについても選択されました。
植物プランクトンの侵入に関するデータ - クロロフィルの総含有量は 1958 年に遡り (Pyrpna、I960)、珪藻および藍藻の培養物からの抽出物を使用した校正を伴う測光法 (Vinberg、Sivko、1953) によって得られました。クロロフィルの初期濃度は、比吸光係数 95 l/g cm に基づいて分光測光法で測定されました (Koski Smith、1948)。1960 年以来、全抽出物中のクロロフィルとカロノイドの個々の形態を決定するために分光測光法が使用されています (Richards、Thompson) 、1952)。 この方法は
この研究は、当時、国内で初めて植物プランクトンの色素の分析に貴重なものであり (Pyrnna、1963)、同時にこの方向での先進的な外国研究 (Humphrey、1963; Tailing、Driver、1963) にも役立ち、その後、植物プランクトンの色素の研究に広く普及しました。 、いくつかの明確化(Parsons、Strickland、1963; SCOR-UNESfCO、1966; Jeffrey、Humphrey、1975)の後、標準として推奨されました(Lorenzen、Jeffrey、1980; Marker et el.、1982; GOST、1990)。
クロロフィル濃度は、1967 年から 1976 年にかけて、Richards と Thompson (1952) の式を使用して 1960 年に計算されました。 - SCOR-UNESCO (1966)、その他の年 - Jeffrey and Humphrey (1975)。 カロテノイドの濃度は、最初の年にはリチャーズとトンプソン (1952) の式を使用して計算され、次にパーソンズとストリックランド (1963) の式を使用して、調査対象の貯水池で優勢だった珪藻プランクトンの日について計算されました。 褐色色素とそれらを除いた純粋なクロロフィル a の濃度は、ローレンツェンの公式 (Lorenzen、1967) を使用して計算されました。
知られているように、Richards と Thompson (1952) の式では、クロロフィル a および b の比吸光係数を過小評価し、クロロフィル c およびカロテノイドの条件値を使用しているため、それらの濃度は後に得られたものとは異なります。 クロロフィル「a」については、この色素のより高い吸光係数 (88 ~ 92 l/g.cm) に基づく他の計算式を使用して計算された値と比較して、25% 過大評価されます。 クロロフィル「b」と「c」の濃度に関するデータは、展開係数とその計算式が改良されるにつれてさらに変化し、150 ~ 200% 変化しました。 カロテノイドの異なる式を使用した計算結果はさらに異なります(最大 2.5 倍); 天然プランクトンにおけるカロテノイドの組成の多様性により、これらの色素の濃度を計算するために適切な吸光係数の値を選択することはほとんど不可能です抽出全体の中で。 したがって、後にクロロフィエートに対するカロテノイドの割合は、抽出物による光吸収へのカロテノイドの寄与が最も大きい領域での消光比によって判断されました - Eva / (Pyrina、Sigareva、1976)、1950年代後半にこれらの目的のために提案されました(バークホルダーら、1959)。 異なる年の計算結果の顕著な偏差を考慮して、植物プランクトン色素レベルのデータの比較は、最も正確に測定されたクロロフィル「a」を使用して実行されました。 同時に、Richards と Thompson の式を使用して得られた初期のデータに 0.75 の補正が導入されました (Pyrina と Elizarova、1975)。
植物プランクトンの色素を測定する方法に取り組む過程で、浸漬油で浄化した後、メンブレンフィルター上に収集した藻類細胞内のクロロフィルを直接測定する非抽出法がテストされました(Yentsch、1957)。 この方法はその単純さで注目を集め、湖のプランクトンの分析で満足のいく結果を示した(Vinberg et al., 1961)。しかし、この方法は湖のプランクトンの分析には受け入れられないことが判明した。
貯留層におけるクロロフィル欠乏は、デトリタスと鉱物懸濁物質の含有量が高いことを特徴とする (Pyrina と Mokeeva、1966)。
3. 一次植物プランクトン生産量の決定
小さな湖の貯水池(プレシーヴォ湖、カーベイ湖)での定常研究では、曝露深さ(有光地帯の5~7層)での光合成を測定するためのサンプリングを伴う古典的な「現場」実験が実施された(ヴィンバーグ、1934年)。貯水池。 遠征船から作業が行われたヴォルガ川カスケードの貯水池と隣接する貯水池では、条件をシミュレートしたデッキインキュベーターでの光合成の初期値を測定するためのサンプル曝露による一次生産量を決定する計算方法が使用されました。水の表層。 同時に、研究の最初の段階では、Yu.I. Sorokin (1958) の修正スキームに従って実験が実行されました。これは、地球のフォトニックゾーンのいくつかの深さのサンプルにおける光合成の強度を測定することを提供します。浸透太陽放射の減衰について得られた値と経験的な関係を持つ貯留層 (Pyrina、1959(a) )、1966)。 その後、彼らは、有光ゾーンの平均サンプルまたは総サンプルに基づいて、保育器(Amshs)内の垂直プロファイルに沿った光合成の最大値を決定することに限定し、その深さの減少は放射線の浸透に基づいて評価されました(Pyrina、 1979年)。 研究対象の深さでの放射線エネルギーは、通常、計算手法を使用したこのような実験で見出されます (Are および Tolstyakov、1969; Pyrina、1989、1993)。 光合成の強度を決定することができない場合は、貯水池の特定の条件に従って以前に確立された値から選択された同化数からAlmを推定するクロロフィル法が使用されました。
光合成の光依存性を研究するための実験は、均一な貯水池の表光帯のいくつかの深さで露出させた自然環境で行われた。底部サンプルは表面から採取されるか、顕著な層状構造の場合は完全に表層内にある。原則として、曝露中はずっと同じ深さで、フォゴインテグレーターのセンサーが作動し、多くの場合、テストサンプルの入ったフラスコがセンサーに取り付けられていました (Pyrina、1967、1974)。水中日射計を使用して一般スペクトルの放射エネルギーの緊急測定が行われ、露光時間中の合計が計算され、その結果、総太陽放射と比較して構築された光合成の一連の光度曲線が得られました。 (Pyrina、Rutkovskaya、1976) 器具を長期間鶏の近くに置くことが不可能だった場合、水中放射量の 1 回限りの測定は、通常は正午、山の麓で、ロックされたサンプルを使用して行われましたか? そして、PARエネルギーが貯留層の表面に到達します。 継続的にrs
海岸または船上で収集されたものは、暴露中に調査深度に到達したことが決定されました。
光合成の強度は、毎日の曝露中の酸素によって評価されました。定常実験では「その場で」夕方に始まり、他の実験では、この方法に関する文献で入手可能な基本的な推奨事項に従って、ステーションでサンプルが採取されたためです(アレキニドル) ., 1973; Vollen weider et al. (1974; Pyrina, 1975,1993)。
4. 研究結果の数学的処理
研究結果に基づいて、植物プランクトンの生産性レベルが異なる貯水池および異なる年で比較されたため、出現した差異の信頼性を確認することが重要であるため、平均値の評価に特別な注意が払われました。得られたデータとその代表性の誤差から (Pyrina, Smetanin, 1982, 1993; Pyrina, Smetanii , Smetanina, 1993)。
貯留層の平均値の決定は、それらが異なる起源の物質に関連しているという事実によって複雑になります。つまり、1 つのサンプルでの繰り返し測定、個々のステーションの異なる深さからのサンプル、貯留層の異なるステーションとセクション、および貯留層の異なる期間です。生育期。 したがって、平均化はいくつかの段階で実行され、それらのいくつかでは、異なる体積の水塊または異なる時間間隔の水塊のデータを平均する場合、平均値は加重平均として計算されました。
貯水池の水域全体のデータを平均する手順には、1 つのサンプルで繰り返し測定した結果に基づいて算術平均を計算し、次に異なる観測点 (または水層) の同じ深さのサンプル データに基づいて算術平均を計算し、次に各セクションの個々の層からのデータに基づく加重算術平均、そして最後に貯留層全体. 最後の 2 つの値は、それぞれ貯留層の層とセクションの体積を考慮して計算されました, でのレベルによって推定されましたそのような評価に必要な深浅地形データが存在しない場合、貯水池の盆地を垂直の壁を持つ物体と同一視することが許可され、その断面の体積の代わりにその面積が計算に入力されました。水層の体積の代わりに、その厚さが計算に入力されました。
成長期の平均データの取得は、個々の観察期間間の時間間隔の算術平均を計算し、次に各期間の日数を考慮して期間全体の加重算術平均を計算することに簡略化されました。 研究対象となった貯水池の成長期は氷のない季節と見なされ、その開始時と終了時の植物プランクトンの量的特性はゼロに等しいとみなされました。 貯水池内の氷の消失と出現の日付は、水文気象局に従って決定されました。
研究された指標のほとんどは直接測定されず、見つからない場合と同様に、適切な式を使用して他の測定値に基づいて計算されるという事実のため
得られた結果から平均値を直接計算することにより、古典的な方法を使用してその代表性誤差を単純に決定することは困難です。 したがって、我々は間接誤差を評価する方法 (Zajdel、1974) を使用しました。この方法では、研究対象の特性がいくつかの変数の関数として考慮されます。 ryを予め定め、その総誤差を微分積分と確率論を用いて各成分の測定誤差を「2次加算」することで求め、測定誤差を迅速に評価できる独自のアルゴリズムとコンピュータプログラムを構築しました。得られた結果を平均します。
Ⅲ. 研究対象の光領域の特徴
aodosg.sha
貯水池が研究された地域は北から南まで広範囲に広がっているため、貯水池の表面に到達する総太陽放射エネルギーの強度は異なります。 平均的な長期データによると、その値はコストロマ地域の年間3500 MJ/m1から5000、アストラハン近郊では夏の真っ盛りの6月にはそれぞれ月600と750 MJ/m1である(ソ連の気候) Reference Book、1966)。ただし、夏期 (7 月から 8 月) の特定の日には、北の貯水池上の太陽エネルギーの強度は同じです!、最大 "4 MDch-"./m2 "" ug pp
水生物学研究では通例であるように (VbiJenweuier et al., 1974)、晴れた日のヴォルガ上流域の水域の一般的なスペクトルの放射線束における PAR の割合は 46% ですが、曇りが増すと 46% になります。 57% (ピルプナ、935 年)。
ヴォルガ盆地の貯水池の深さにおける全太陽放射の分布を観察したところ、それらの最大透明度(セッキ円盤に沿って最大2メートル)は全体の1%であることが示されました。
水中に放出された放射線量は2メートル以下の深さで記録されていますが、水の透明度が2.4メートルまでのヴォルゴグラード貯水池でのみ、そのような放射線値は深さ3〜4メートルで記録されました(Rutkovskaya、1965)。 より透明な湖水では、水に入る放射線の約 1% がさらに深いところで観察されます。ラドガ湖では水深 5 ~ 6 m (モキエフスキー、1968)。 オネガでは 6 ~ 7 m (モキエフスキー、1969; ピリナ、1975(a))。 湖の深さは6〜8メートルです。 プレシチェエヴォ (ピリナ、1989(a))。 日中および南に向かうにつれて地平線上の太陽の高さが増加するにつれて、放射線の浸透深さの増加も認められました(Rutkovskaya、1965)。
水に入ると、最も外側の長波線と短波線が最初に減衰し、PAR に近い放射線だけが 1 メートルより深く浸透します (Rutkovskaya, 1965; Pyrina, 1965; Pokatilova, 1993)。 貯水池内の太陽放射を減衰させる主な役割は、デトリタスと鉱物懸濁物によって演じられます。 このような背景から、植物プランクトンによる光の吸収、いわゆる「自己シェーディング効果」(Talling、1960) は弱く感じられます。 これは、水塊の安定性が高まる時期と一致し、水が青緑色で「咲く」夏にのみ観察でき、懸濁粒子の総量に占める藻類の割合が優勢になります(Pyrina、Rutkovskaya、イリンスキー、1972)。 そして湖などの透明な湖水に限ります。 Pleshcheyev によれば、植物プランクトンの光学的影響は、ホサーモミーの期間中であっても非常に明確に追跡することができます (Pyrina、Sigareva、Balonov、1989)。
貯水池の濁った水では、深部から戻る拡散放射線の値の増加が観察されます。 ルイビンスク貯水池でのPAR測定によると、Secnディスク全体の透明度が0.7〜1.5メートルで、目は水に入る放射線の2〜10%に相当しましたが、オネガ湖では透明度が約4メートルで、それよりも低いです。 1% (ピリナ、1975(a))。
ルイビンスク貯水池とオネガ湖での透過性PARのスペクトル測定(図1)は、水の表層には青色の光だけでなく、600 nmを超える波長の赤オレンジ色の光線がかなりの割合で存在することを示しました( X - 380-480 nm)。 ただし、どちらもすぐに消え、緑色の光線が最も深く浸透します (X = 480600 nm)。 プラチナコバルトスケールで色値が 70 度を超える水では、水の最上層によって吸収される青色放射線 (X = 380480 nm) の割合の減少が顕著であることが認められました (Pyrina、1975(a))。 。
冬の観察。 ルイビンスク貯水池で実施された雪と積雪を通るPARの浸透に関する研究では、積雪20cm、氷の厚さ80cmの典型的な条件下では、入射放射線のエネルギーが0.04%以下に達するに過ぎないことが示された。水 (200 ~ 250 W/m に相当)。これは、雪からの強い反射 (80%)、雪からの後方散乱の割合 (3%)、および雪によって遅延された放射 (13%) によるものです。雪が溶けた後、氷の下に浸透する放射線量は18〜20%に増加します。
米。 I. オネガ湖 (a ~ c) およびルイビンスク貯水池 (d ~ f) の水柱へのさまざまな波長の太陽光線の浸透、% o g
入ってくる放射線エネルギー。
1 - L = 380-800 μm; 2-X-480-800nm; 3 - L = 600-800 k\<; 4- Л = 680-80") им; вертикаль вниз - прозрачность по белому диску; цифры гмд пен ■ цветность по пяатиново-кобалътовой шкале.
そしてそれは 2 メガの深さ (0.4%) まで追跡できます。 同時に、植物プランクトンの繁殖も注目されています (Pyrina、1984.1985(a))。
特別な装置を必要とする水中PAR測定の複雑さと、比較的小さな水層を通過した後は太陽スペクトルのこの部分のみが残るという事実を考慮して、積分放射束の減衰パターンを研究しました。水文気象サービスの標準的な光量測定データを使用するために貯水池に保管されます。
太陽放射の経路を詳細に記述する場合、古典的なブーゲーの法則が広く使用されていますが、知られているように、この法則は単色放射の場合にのみ有効であるため、完全に正当なものではありません。 ブーゲーの公式を使用して計算された放射線値:
(ここで、1rと10は深さrで水に入った*放射線の強度、および「垂直減衰の指標」です)は、測定値から逸脱します - 水の上層ではそれらは過大評価され、深層ではそれらは過大評価されます。は過小評価されており、このことは SDovgy, 1977; Scumec, 1975; Kirk, 1983 の多くの研究で指摘されています)。この偏差を補償するために、式 (1) の指数にパラメータ r の累乗を導入することが提案されました。 1 未満、特に 0.5 に等しい (Rosenberg、1967) および自然データ (Are、Tolstyakov、1969; Larin、1973) は、この修正を加えて透過放射線を計算できる可能性を確認しました。
しかし、本研究における修正式(2)の適用により、下層地平線で計算された放射線強度の大幅な過大評価が明らかになった。これは、光合成の深さと1以下の水柱におけるその積分値を決定する際に考慮する必要がある。 m1。 この現象の研究により、深さ方向の太陽放射の経路の最も適切な記述は、パラメーター r の指数 (u) の可変値を使用して式 (2) を使用することによって得られることが示されました。
nの値は水の色によって異なり、低い値では理論値に近くなります。 たとえば、湖の場合。 10度「=0.6」のプラチナコバルトスケールの水彩画のプレシチェエヴォ、および
ルイビンスク貯水池、色は 50 度 l = 0.8 (Pyrina、1989)。
したがって、調査対象の貯水池における水中光合成の光条件の研究では、太陽放射の浸透深さが広範囲に渡っており、強い日射量ではヴォルガ貯水池の透明度の低い水域では 2 ~ 5 m から 7 ~ 12 m まで変化することが示されました。オネガ湖とプレシチェエヴォ湖にいます。 上部内にある比較的浅い深さでの極短波線と長波線が急速に減衰するためです。 0.5 メートルの層では、透過放射線は PAR スペクトルで同一になります。 水による選択的減衰により、その深さの進行が指数関数則から逸脱することが観察されますが、これは数学的に補正することができます。 「これにより、水文学研究において十分な精度で、最も近い光量測定ステーションによって陸上で記録されたものを含む、貯水池上の太陽放射の強度から光合成に必要な太陽放射のエネルギーを計算することが可能になります。
1958 年にイワンコボ、ルイビンスク、クイビシェフ貯水池で開始され、その後調査対象となったすべての水域で実施された植物プランクトン色素の測定では、その濃度に大きな変動が見られました (表 1)。 主な色素であるクロロフィル「a」の場合、それらは1リットルあたり100分の1マイクログラムから00以上までの範囲にあります。「貯水池の有光ゾーンで計算されます。ただし、各シュノロエモ」は、一定レベルの平均値によって特徴付けられます。ベタインの季節と最大達成値に基づいて、その水域の栄養学的関連性を判断することができます。この原則によれば、単栄養性のオネガ湖から富栄養性のイワンコッホまで、ほぼすべての水域の貯水池が相続によって覆われました。貯水池。
貯水池内のクロロフィル濃度の変化は主に植物プランクトンの季節動態に関連しており、その最大値はその大量成長の期間中(原則として5月と6月に観察されます)に観察されます。 ] ルイビンスク貯水池の 1a nrig^p^ によれば、特定の時期におけるこれらの最大値の高さとタイミングは、その年の水文気象学的特徴に依存することが示されています (Pyrina、Sigarepa、1986)。 ここでは、0.2 μg/l 以下のクロロフィルが送達されたときの、氷下の gter.tod におけるクロロフィルの uro-ttagt、tsoggceggtrpcpy を追跡しました。 雪が消え、光合成活性放射線が氷の下に約 1 MJ/m2 ■ 日浸透した後にのみ、クロロフィル濃度は 0.4 μg/l に上昇し、植物プランクトンの植生が始まりました (Pyrina, 1985(a))。
表 1
貯留年数、クロロフィル、供給源
月 µg/l 情報
イワンコフスコエ 1958、V-IX 12.5 ピリナ、1966
vdhr。 0 - 2 m 1970、V-X 13.3 エリザロワ、1976
1973年から1974年、V-X。 26.7-31.8 ピリナ、シガレバ、
1978 年、V-X 14.2 ピリナ、シガレバ、
■未公開
ルイビンスコエ 1958、V-X 6.6 ピリナ、1966
メインリーチ。 1969-1971、V-X 3.4 - 6.7 エリザロワ、1973、
1972-1976、V-X 6.2-10.0 ピリナ、シガレバ、
1977-1979、V-X 6.6 - Yu.o Mineeva、Pyrina、
1980-1982、V-X 9.3-18.2 ピリナ、ミネバ、
1983-1985、V-X 15.4-19.2 ピリナ、1991
1986 ~ 1990 年、V-X 9.4 ~ 13.8 同上。
1991 ~ 1993 年、V-X 12.6 ~ .14.6 ピリナ、未発表。
クイビシェフスコエ 1958、VI-X 7.9 ピリナ、1966
vdhr。 0~3m
ホワイトレイク。 1976-1977、V-X 3.8 -5.0 ピリナ、ミネバ、
0~2メートル博士、1981年
オズ。 プレシチェエヴォ 1983-1985、V-X 6.2-10.0 ピリナ、シガレバ、
バロノフ、1989 年
オズ。 B. カーベイ 1969、VII-VIH 2.0 - エリザロワ、ピリ-
オン、ゲッツェン、1976
オネガ 1967-1968、0.57 - 0.95 ピリナ、エリザロワ、
湖。 0 - 5 m VII-VIIÍ ニコラエフ、1973
ラドガ湖 - 1973-1974、VIII 4.60 ピリナ、トリフォ-
ろ。 0 - 4、0 - 5 m 新星、1979 年
R. エニセイ。 1984-1985、4.2-7.2 ピリナ、プリマ-
0 m - 最下部 VWX チェンコ、1993
注: データは夏季の平均値であるオネガ湖とラドガ湖を除いて、貯水池と生育期の平均値が示されています。
水域全体のクロロフィル濃度の違いはそれほど顕著ではありません。 栄養素が豊富な川の水が流入する地域、および浅瀬、特に孤立していて大型植物が弱く生い茂る地域のみ、腐敗の含有量が増加します(Egshzarova、1976、1978; Elizarova、Sigareva、1976; Elizarova、Pyrina、Getsen、1976) ; ピリナ、1978; ピリナ、プルタロワ、ニコラエフ、1973; ピリナ、プリマチェンコ、1993)。 均質な水塊内の小規模な差異は比較的小さく、主に動的な要因の作用、つまり植物プランクトンの総合的な輸送、風の混合、水の層化を決定する循環流と関連しています (Pyrina、Sigareva、Balonov、1989; Pyrina、 Mineeva、Sigareva 他、1993)。
研究されたほとんどの貯留層では、クロロフィルの変換生成物である葉緑素がかなりの量で認められました。 たとえば、ルイビンスク貯水池では、褐色色素コシアシの量は純粋なクロロフィルの量の 20 ~ 30% ですが、初夏の植物プランクトンの減少期には、特に水の下層で 60 ~ 70% に上昇します (Pyrina) 、Sigareva、1986;Pyrina、Mineeva、1990)。 現時点のプレシチェエヴォ湖では、表層 2 メートルの層で同量の褐色色素が観察されており (Pyrina、Sigareva、Balonov、1989; Pyrina、1992)、それに加えて草食性動物プランクトンの量が増加しています (Stolbunova、1989)。 。 通常、青緑色の水の「開花」に関連する夏の最大温暖化の期間中にのみ、褐色色素の含有量は10%未満になります。 これは、太陽放射の集中的な摂取と一致しており、太陽の破壊的な影響により照射された領域にクロロフィルを含むクロロフィルが蓄積するのを防ぎます (Yentsch, 1965; Moreth Yetisei), 1970)。このプロセスが起こる水の上層の安定性 (Pyrina, Sigareva. 19X6)。
他の緑色色素であるクロロフィル「b」と「c」は、はるかに少量で発見されました。 南方珪藻、隠蔽植物、および二葉植物のクロロフィル c" の含有量は、クロロフィル "b" (数パーセント) よりも総クロロフィルのわずかに高い (最大 30%)。ただし、沿岸、河口、その他の水域は栄養分が豊富で、したがって、緑藻およびユーグレナ藻 - クロロフィル「b」のキャリア、その含有量は10%に達する可能性があります。
任意の単位で測定されるカロテノイドは、通常、量的にはクロロフィルに対応します。 水の生産性が低下するにつれて、絶対含有量と消光比の両方において、クロロフィルとの比率が増加する傾向が見られました。 たとえば、ルイビンスク貯水池の中央部では河川流域や沿岸地域よりも高く、この貯水池の中央部ではイワンコヴォ貯水池よりも高い(Elizarova、1973、976; Elizarova、シガレバ、1976; ピリナ、シガレバ
1978年、1986年)。 オネガ湖、特にその深部、およびツンドラ地帯のハルベイ湖ではさらに高くなります (ピリナ、エリザロワ、ニコラエフ、1973 年; エリザロワ、ピリナ、ゲッセン、1976 年)。 例外はエニセイ川で、水の生産性が比較的低く、この比率が低いという特徴があります (Pyrina、Priymachenko、1993)。 季節の側面で緑色と黄色の色素の比率を分析すると、植物プランクトンの増加が初夏の最小期間、特に植物ファージの繁殖中に観察され (Pyrina、Sigareva、Balonov、1989)、冬には非常に大幅な増加が観察されます (Pyrina) 、1985/a)。
植物プランクトンバイオマス中のクロロフィル「a」(通常 2 ~ 5 μg/mg)およびカロテノイド(1 ~ 5 μBri/mg)の比含有量に関しては、研究された水域で有意な差は検出されません(Pyrina、1963;エリザロワ、1974、1976; ピリナ、エリザロワ、1975; エリザロワ、ピリナ、ゲッツェン、1976; ピリナ、プリマチェンコ、1993)。 これらの藻類の培養物 (Pyrina、Elizarova、1971) のように、顕著な割合で緑藻類を含む植物プランクトンのバイオマスでは、クロロフィル含有量の増加のみが認められました。 Biriapsis 属の細胞種 (Pyrina、Sigareva、Balonov、1989)、
したがって、植物プランクトン色素の季節的および空間的動態の一般的なパターンは類似していますが、調査対象の水域におけるそれらの濃度レベルは大きく異なり、非生産的なオネガ湖とオネガ湖、およびオネガ湖とオネガ湖での成長期には平均して 1 ~ 2 μg/l の範囲です。ボリシャヤ・ハーベイは30μg/lに - 富栄養化したイワンコフスキー貯水池で? しかし、植物プランクトンのバイオマス中のクロロフィルの具体的な含有量は、色素のセットと主要な種の組成が類似しており、非常に近いです。 光学的に異なる貯留層からの植物プランクトンの光適応に関連する色素特性の特殊性の痕跡はなく、光条件の役割は、期間中の貯留層の受光層におけるクロロフィル誘導体の形成と蓄積のプロセスを制御することにおいてのみ明らかにされます。高気圧型の安定した天気。
V、植物プランクトンの光合成活性
1 m2 未満の水柱における植物プランクトンの一次生産量に基づくと、調査対象の水域のほとんどは中栄養のカテゴリーに分類されますが、このタイプの水の栄養レベルはさまざまです。ツンドラの貧栄養に近いレベルからB. ハーベイ湖からルイビンスク貯水池の富栄養との境界まで。 オネガ湖のみが典型的な貧栄養貯水池として分類されており、イワンコフスコエ貯水池とチェボクサル地区は典型的な富栄養貯水池として分類されています。 湖は特別な場所を占めています。 プレシチェエヴォ、そのために
表2
成長期中の調査対象の水域における植物プランクトンの一次生産量の平均値(g C/m2 日)
イワンコフスコエ 1958 0.90 ピリナ、1966
vdhr。 1970 ~ 1973 年 0.90 ~ 1.28 ピリナ。 1978年
1974-1976 0.36 - 0.68 サッポ、1981
1979 ~ 1980 年 0.91 ~ 1.35 タラセンコ、1983 年。
ルイビンスク貯水池 1955 0.28 ソローキン、1958
1958 0.54 ピリナ、1966
1959 0.59 ロマネンコ、1466
1001-1905 0.10-0.19 ロマネンコ、1985
1966-1971 0.21-0.48 同じ
1972-1973 0.69 - 0.72 同じ
1974 ~ 1980 年 0.18 ~ 0.35 同じ
1981 0.83 同
1982 0.52 ミネバ、1990
ゴルコフスコエ 1956 0.41 ソローキン、ロザノワ、
vdhr。 ソコロワ、1959 年
1967 0.76 タラソワ、1973
1972年 0.89タラソワ。 1977年
1974-1979 0.31 - 0.85 シュメレフ、スボプシャ。 1983年
R. ヴォルガ 一部 - 1966 2.25 タラソワ、1970
チェボクサル-
クイビシェフスコエ 1957 0.66 サルマノフ、ソローキン、
1958 0.83 ピリナ、1966
1965-1966 0.37 - 0.72 イヴァチン、1968、1970
1967-1971 0.32 - 0.56 イバチン、1974、1983
サラトフ 1971-1973 0.32-0.50 ジュバン、1975、1976、
ヴォルゴグラード 1965-1968 0.30 - 0.63 ダレチナ、1971、1976
vdhr。 1969-1974 0.33 -0.64 ダレチナ、ゲラシモ -
1975 1.50 同じ
表 2 の続き
貯水池 年 主な情報源
研究生産情報
ホワイト レイク 1976-1977 0.21 -0.31 ピリナ、ミネバ、および
オズ。 プレシチェエヴォ 1983-1984 1.36- 1.86 ピリナ、シガレバ、
ジュバン、)989
オネガ湖 1966、VII 0.05 - 0.20 ソローキン、フェドロフ、
ラドガ湖 1973-1974、0.32-0.60 ピリナ、トリフォノバ、
オズ。 B、カーベイ。 1968-1969 0.14-0.26 ピリナ、ゲッセン、
ヴァインシュテイ、1976年、
R. エニセイ 1984 0.86 プリンマチェンコ、1993
注: 炭素に変換する場合、酸素法のデータは 0.375 倍されます。 オネガ湖とラドガ湖の場合、最大値は開いた部分に与えられます
ここではクロロフィル含有量と水の単位体積あたりの光合成の強度が比較的低いという事実にもかかわらず、一次生産量の最高値が得られました(夏には最大3 g Og/m2日)。
研究対象となった水域におけるプランクトンの光合成の季節経過は一般的なパターンによって特徴づけられ、主に藻類の量の変化によって決定されます。 ルイビンスクやクイビシェフなどの湖や湖に似た貯水池では、主要な藻類グループの変化の周期性がはっきりと見えるため、春と夏のピークが観察されますが、通常、最初のピークは限定的な影響によりそれほど顕著ではありません。 (Pyrina, 1966; Pyrina, Mineeva, and al., 1981; Pyrina, Sigareva, Dzyuban, 1989)、流量が増加した河川タイプの貯水池、たとえば、成長期を通じて珪藻が蔓延するイワンコフスキーでは、光合成の季節過程がよりスムーズになります。 しかし、ここでは真夏でもその最大値が観察されます(Pyrina、1978)。 浅海では、季節動態の一般的な性質は深海域と似ていますが、光合成のピークが初期の時期にシフトしていることが注目されました (Pyrina、Bashkatova、Sigareva、1976)。 ルイビンスク貯水池での個々の観察 (Romanenko、1971) から判断すると、冬には、雪と氷に覆われた下の光不足のため、光合成は非常に弱くなっています (Pyrina、1982、1985(a))。
一次生産における空間的な違いは、植物プランクトンの発達の程度(クロロフィル含有量)と密接に関係しています。 ただし、これは水の単位体積あたりの光合成の強度の観点からのみ明確に見えます。 貯水池の単位面積ごとに計算されたその積分値では、このパターンは違反されており、これは何らかの形で光環境の条件に関連しているため、受光層より浅い深さの植物プランクトンが豊富な浅瀬では、 、ヨウ素の一次生産量 1m: 底部の感光層が制限されているため、比較的少量です (Pyrina、Bashkatova、Sigareva、1976; Pyrina、1978; Pyrina、Sigareva、Dzyuban、IW)。強く「咲く」青緑色で水域では、ヨウ素は下層の植物プランクトンへの光エネルギーのアクセスにおける「自己シェーディング効果」によって制限され、ペットの光合成若さが減少します。このような水域は、顕著な光合成の独特の垂直プロファイルによって特徴付けられます。逆に、透明なプレシチェエヴォ湖やヴォルガ川下流域の一部の地域など、太陽光線が深く浸透すると、さらに太陽エネルギーの量が増加し、一次エネルギーが増加します。 1 m3 未満の水柱での生産は、藻類のバイオマスが比較的小さい場合に高レベルに達します (Pyrina、Rutkovskaya、1976; ゲラシモワ、1981年。 ピリナ、シガレバ、ジュバン、1989)。 湖で プレシチェエボでは、単位面積当たりの一次生産量の増加は、最適な条件を求めて活発に混合することができる、夏の植物プランクトン(Ceraium 属のディノフト藻類)の大量形態の生態学的特徴によるものでもある(Heaney、Talling、1980 年) )。 強烈な太陽放射の下では、それらは最大光合成ゾーンの特定の深さに蓄積し、1 mあたり水100リットルあたりのヨウ素の値の増加に寄与します。 (ピリナ、シガレバ、ジュバン、14S9)。
垂直方向のプロファイルに沿った光同期性強度の分布パターンは、ドイツのすべての研究で一般的に類似しており、主に特定の深さでのパージェションの量によって決定されます。 マキシマムフォユエシュペ! 放射照度が約 4.0 MD-k/m2 日 PAR の深さで観察され、その減衰は 0.02 MJ/u3 ■ 日です。 晴れた日には、そのような条件は、それぞれ、t.i Pleshcheyevまでの深さ1〜2 mおよび10 mで作成されます(「Pyrina、Sigareva。Dzyuban、1989)。1.0〜1 5 n 1〜9 m i、オンチ湖(Sorokin 。ヒョードル ")"、iУ69>、Ö-0.5、3〜6 m、L. Kh湖。<(" vi* (üupiuu, ;сцсн, Ьакншчсин, 1976), 0-0,4 и 2-5 м в волжских водохранилищах (Пырина, Рутковская, 1976). Так, практически одна и та же энергия облученности (96 кал или 4,02 МДж на I м2 в сутки) наблюдалась на глубинах максимального фотосинтеза в Рыбинском вопохпл-ннлнще как весной, при интенсивной вегетации диатомовых, так и ж-(ом, когда в массе развивались сине зеленые (рис. 2). Однако в одном и том же водоеме эти, соответствующие характерным параметрам фото-синтезнрующей зоны, глубины могут значительно варьировать в шви-
米。 2. 日射の垂直分布 (X = 380-800 nm) と珪藻または雪緑色藻類が優勢な植物プランクトンの光合成。 ルイビンスク貯水池、1972年
a - 珪藻の「開花」(5 月 16 日)、b - 相乗効果の「開花」(7 月 7 日)。 1 - 光合成 (mg O/l.day)、2 - 放射線 e MJ/m "スープを MRE スケールに換算したもの (方法論的レター... 982、引用元: Evnevich、1984)。矢印は、放射線のエネルギーを示します。最大光合成の深さ。
太陽放射の流入からのシミュレーション。これは、1 mg未満の一次生産量の積分値の変化につながります(Pyrina、Mineeva、印刷中)。
研究された貯水池における植物プランクトンの光合成中の太陽エネルギー利用効率は、水に入射する放射線の統合スペクトルの0.01〜1%、またはPARエネルギーの0.022%の範囲にあります。 最高値(約1%)は、豊富な植物プランクトンを特徴とするチェボクサル貯水池、クイビシェフ貯水池とイワンコフスキー貯水池(Pyrina、1967(a)、1978)、および湖の地域で得られました。 Pleshcheev - 光合成ゾーンの深さによる(Pyrina、Sigareva、Dzyuban、1989)。 これらの値は、生産性の高い淡水として知られている最大値に相当します (Talling et al., 1973)。 湖の例を使用する プレシチェエヴォは、太陽光利用の過程における光環境の指標と植物プランクトンの量の相互影響を追跡しました。植物プランクトンの量は、クロロフィルの濃度または10μg/lに比例して増加し、さらに増加すると減少し、「セルフシェーディング効果」(Pyrina、Sigareva、Dzyuban、1989)。
鉛直断面図に沿った最大光合成強度と、植物プランクトンの同化活動を特徴づける藻類または藻類に含まれるクロロフィルのバイオマスとの関係は、すべての水域でかなり狭い範囲内で変化します。 ほとんどの場合、光合成のバイオマスに対する比率 (バイオマス活動係数) は 0.3 ~ 0.4 mg Qi/mg、クロロフィル (同化数) に対する比率は 1 日あたり 0.10 ~ 0.20 mg Og/μg です。 それらの値の増加は、河口、浅瀬、その他の地域で典型的であり、植物プランクトンでは緑藻がかなりの割合を占めています (Pyrina, 1959 (a), ¡967 (5), 1978; Pyrina. Trifonova.」 ." 79; Pyrina et al. 1981); シガレバ、1984年。 ピリーナ、スンギャロイ、バロイフ、1989)。
一次生産物の対象を絞った研究では、垂直方向のプロファイルに沿ったその最大値は、浮遊プランクトンとそれに含まれるクロロフィルの量によって決定され、柱の積分値は光領域の特性によって決定されることが示されています。貯水池内での光合成の広がりの深さを決定します。その結果、淡い色の透明な水では、吸収されたほとんどない放射エネルギーを受け取り、クロロフィルと入射太陽放射の単位あたりの光合成の収量が送信されます。光係数が有意な差を決定します。地平線上の太陽の高さに応じて、強力な太陽エネルギーの摂取期間とその浸透の深さに関連する一次生産の年間平均値。
VI. 植物の光合成の光依存性のオセイカ-
水中の太陽放射のエネルギーに応じて植物プランクトンの光合成活動を研究する実験は、ルイビンスク貯水池と湖のヴォルガ中下流の広い水域で実施されました。 Pleshcheev により、光合成の一連の典型的な光度曲線が得られました。 「正規化」後、各サンプルの最大値を 100% として、同様の照射条件下で確立されたさまざまな実験からのデータは、PAR エネルギーと一般的な太陽スペクトルの両方に関して、非常に密接にグループ化されていることが判明しました (図 3) それらの不一致の一部のみが、主要な藻類グループの組成に依存しています: 緑色が優勢な植物プランクトンの場合、点は高い照射エネルギーの方向にシフトし、珪藻由来の植物プランクトン、特に青緑色の植物プランクトンの場合は、点がシフトしました。藻類 - より低い値の領域へ(Pyrina、1967; Pyrina、Rutkovskaya、1976)。
このような結果は、海棚における底生藻類の垂直分布のパターンと一致しており、緑色の代表的な藻類は浅い深さの明るい条件下でより頻繁に成長するのに対し、珪藻、茶色、特に赤色の藻類はより深くまで浸透します。 同様の現象がいくつかの深層淡水域でも観察されている (Gessner, 1955; Kirk, 1983)。 色素の光吸収能力に関連する光順応の兆候は、さまざまな深さから飼育された海洋植物プランクトンでも観察されました (Koblenz-Mishke、1980)。 しかし、ルイビンスク貯水池からのプランクトンの分離(ピリナ、1959年)を含む培養物を用いた実験(カーティス、ジュデイ、1937年;市村、有賀、1958年など)では、SVSTに対する藻類の反応に同様のパターンは見られなかった。追跡された顔料のセットに応じて。
この矛盾と、ヴォルガ川の植物プランクトンは、研究された他の貯水池と同様に、主に珪藻とスノーグリーンによって表され、光との関係が非常に近いことが判明したという事実を考慮に入れて、単一の光度曲線を作成します。ヴォルガ貯水池で行われたすべての実験の結果を総合して構築されました(図3)。 この場合、幅広い研究者が最もアクセスしやすい、一般的なスペクトルのエネルギーに関するデータを使用しました。 おおよそ、つまり ヴォルガ貯水池では比較的ほとんど発現していない光阻害を考慮せずに、水中に浸透する太陽放射に対する植物プランクトンの光合成の依存性は、次の形式の方程式で記述されます。
Az"=Iz/(0.35 + 0.009 l.J, (4)
5 10 50 yua)50 ¿50 /Gal!cis
1 日あたりのアイビー放射線量
米。 3. 異なる強度の日射入力におけるヴォルガ貯水池における植物プランクトンの光合成の相対速度(一般スペクトル)。
1,2 - それぞれ藍藻類または珪藻が優勢な植物プランクトン。 3 - 緑色の植物プランクトンが優勢な混合植物プランクトン。垂直方向のプロファイルに沿った最大値の % での光合成。 太陽放射エネルギー (MD/s/m 小屋単位、MRE スケール)。
ここで、1gおよびAz""の放射線エネルギーと深さrにおける光合成の強度(垂直プロファイルに沿った最大値の%)。 Ar -(AJAMaKC) 100。Ar と Amax は絶対単位です。 結果として得られる方程式 (4) は、よく知られているミカエリス・メンテン方程式に似ており、光合成に対する光の影響を説明するための一般的なオプションの中で最も単純なものと考えることができます。 特に、これはバニスター方程式 (Bannister、1979) の変形です。
A,=AMaKC!/(Ikm+Im) Ш (5)
(ここで、A j と Amax は、それぞれ放射エネルギー l および垂直プロファイルに沿った最大値における光合成の強度です。/^ ■ 定数)、これは非常に普遍的であり、t -I ではミカエリス・メンテン方程式に変わり、t では= 2 を式に代入します。Smith (Brother、1994)。 後者は、知られているように、1 mg の水柱における積分一次生産量を計算するために採用された Talling モデルの基礎を形成します (Tailing, 1957; Vollenweider, et al., 1974)。 パラメータ m の値を変えることにより、より高い光強度 (たとえば、m - 2 の場合) または低い光強度 (>>u = 3) での飽和領域を持つ光曲線を記述することができます。
したがって、導出された式 (4) は、かなり平坦な傾きを持つ光合成の光度曲線に対応し、最大値が照射量が増加した領域にシフトし、光阻害が弱く発現します。 この種の依存性は、深さとともに減少するにつれてスペクトル的に変化する、実験における放射照度の特殊性にある程度関連している可能性があります。 それでもなお、水によって弱められた光に対する顕著な適応の欠如は、明らかに、研究対象の水域における植物プランクトンの大部分の特徴であると考えることができ、そこでは混合因子が重要であり、藻類細胞が深層水層に長期間留まる必要がない時間。
VII. 植物プランクトンの生産性の年々変動における、異なる年の光体制の特徴の役割
クロロフィル含有量に関する最も完全な一連のデータが得られたルイビンスク貯水池の例を使用して、人為的要因と気候的要因の役割を区別して評価することで、長期計画における植物プランクトンの生産性レベルの変化が検討されました。特に、太陽放射体制 (Pyrina、1991)。
これらのデータの分析により、氷のない期間中の貯水池内の平均クロロフィル含有量が 197 から増加していることが示されました。 g. (図 4)。 80 年代までに、それは臨界レベル (10 μg/l) に達し、それを超えると動物動物は通常富栄養として分類されます。 これは栄養素、特に総リンの量の増加と一致しており、主貯水池の濃度は 1965 年と 1970 年に 33 ~ 47 µg/l から増加しました。 (個々の季節の平均で) 1980 年には最大 40 ~ 60 µg/l (Bylinkina、Trifonova、1978; Razgulin、Gapeeva、Litvinov、1982)。 貯水池のこの部分のクロロフィル濃度が特に急激に増加していることからわかるように、それらは主にチェレポヴェツ工業団地からの廃水を受け取るシェクスニンスキー流域を通って侵入する。
一般的な増加傾向を背景に、クロロフィル濃度の高い上昇が目立ったが、これは高気圧性気候の年 (1972 年、198!、1984 年) の植物プランクトンの発生に対応しており、これは太陽放射の強烈な流入によるものである: 15 -16 MJ/m* 日。 生育期(5 月から 10 月)の平均で、夏期の最も頻繁な値は 20-25 MD/m2 ■ 日の範囲です。 その後、色素含有量はわずかに減少しましたが、フレア前の数年間よりも高いままであり、地球に入る太陽放射エネルギーが増加する次の期間が始まるまで、数年間はこのレベル付近に留まりました(図4)。 近くのイワンコフスコエ貯水池と湖。 これらの年(1972年、1973年、1984年)のプレシチェエボでは、クロロフィル含有量(表2)および一次植物プランクトン生産量(表3)の値の増加も観察されました。 一次生産レベルの増加は、水文気象条件と同様の年、たとえば 1975 年に他のヴォルガ貯水池でも発生しました (表 3)。 湖で Windermnr は、春の珪藻のピークバイオマスの形成に対する太陽放射のプラスの影響を明らかにしました (K"ea! e1 a!., 1991)。これは本当です (.kiyu は、植物プランクトンバイオマス (クロロフィル含有量) の年間平均レベルに影響を与えました。これは、同時期の他の要因によって主に決定された(TaShiv、1993)。
高気圧性気候の年における植物プランクトンの生産性の大幅な増加は、光合成と藻類バイオマスの成長に必要な一連の複雑な条件の影響によって説明されます。 太陽放射の激しい摂取により、光合成の広がりの深さが増加し、それに応じて貯水池の単位面積あたりの水柱の積分値が増加します。 そのような年に特徴的な風が弱まった条件では、水柱の加熱が増加し、その安定性が増加します。 これにより、活性(誘導体を含まない)クロロフィルを多く含む藻類が照射された領域に長期間滞留し、その光合成と成長が促進されます。 生物起源の要素、塵によって両方のプロセスを刺激することが可能です
17 16 15 N 13 12 I 10 9 V 7 6
1958 1971. 1975 1979 1983 1987 1991
1969 197E 1977 1901 1985 1989 1993 ガーナ
図4. ルイビンスク貯水池における入射放射線の強度とクロロフィル含有量の長期変化
太陽
/ - ルイビンスク水文気象観測所による、5月から10月の平均の総太陽放射エネルギー(総スペクトル)、MJ/m1日。 2 主貯水池範囲の標準観測所での観測による、氷のない期間中の水の上層 2 メートルにおけるクロロフィル「a」(色素色素を含む)の平均含有量、μg/l。
細菌による有機物の破壊によって破壊され、熱によっても破壊されます(Romanenko、1985)。 明らかに、太陽放射が激しい期間中、貯水池の生態系は光合成中に利用されるエネルギーで豊かになり、植物プランクトンの生命プロセスは実際に栄養素によって刺激されます。 バイオマスの成長を確実にします。
得られた結果は、水システムが機能するための主な条件、つまりそれを維持するための太陽エネルギーの強制的な流入を明らかにしました。 この観点から見ると、光は、浮遊性フィトセノーゼの生産性と、その富栄養化のプロセスを含む貯留層全体の制御において、生物起源の要素と同じくらい重要な役割を果たしている。 したがって、この世界的な問題の解決策は、ライト スターの影響を評価することに関連しています。
結論
主な研究結果は以下のとおりです。
1. 水の透明度に応じて、調査対象の貯水池における太陽光の浸透深さは大きく異なり、ヴォルガ貯水池の 2 ~ 5 m から、強い日射量下でのオネガ湖とプレシチェエヴォ湖の 7 ~ 12 m までの範囲です。 「約 0.5 ~ 1.5 m の浅い深さのコーラの層の中で極度の短波と長波の光線が減衰するため、透過放射線は PAR のスペクトル内で同一になります。PAR 内ではより深く広がっています」スペクトルの青色領域の「光線」は、特に底部の色が増加すると最も集中的に減衰され、次に赤色になり、一方、緑色は最大深度まで浸透します。放射線の選択的減衰を伴う「イーンツ」では、その深さコードが逸脱します。この偏差は数学的に補正することができ、最寄りのアクトムストレンチェスコン観測点で登録されたものを含む、水域全体の太陽放射の強度から水中光合成に必要な照射エネルギーを計算することが可能になります。
2. 植物の主要な色素であるクロロフィル「a」の含有量は、オネガ湖とツンドラ湖の非生産的な水域では、飼育期間中平均 1 ~ 2 μg/db b の範囲で変化します。 B. 富栄養化したイワンコボ貯水池では最大 30 μg/l のカーベイがあり、植物プランクトンの大量増殖期中の最大値はそれぞれ 1.5-3 μg/l から 100 以上です。 色素含有量のレベルが異なる水域の各グループにおけるクロロフィル含有量の季節動態の独自性を追跡することができます。つまり、湖では鋭いピークと比較的弱く表現された夏のピークです。 プレシチー
大きさは同じ - ホワイト湖、春、夏、秋(または夏秋) - ルイビンスク貯水池、ある夏 - イワンコフスコエ貯水池、およびツンドラのハルベイ湖、そして明らかにオネガ湖。 冬には、厚い雪と氷の層の下で光がほぼ完全になくなるため、クロロフィル濃度は 0 に近くなります。すべての水域におけるクロロフィル含有量の空間変動パターンは非常に類似しており、主に次の条件に関連しています。流体力学的体制。 クロロフィルの特定の含有量、および異なる水域からの植物プランクトンの主要な藻類の均一な組成に関する他の色素との比、さらには光学的に異なる水域におけるクロロフィルの色素系の光適応の兆候には有意な差は見出されませんでした。そして季節的な面でも。 同時に、クロロフィル誘導体の形成と蓄積の過程における太陽放射の制限的な役割が明らかになり、そのため、高気圧性の天候で光合成に最も好ましい期間中、アクティブファンドの高い含有量が維持されます。 このような時期が優勢な年には、貯水池内のクロロフィル含有量の全体的なレベルが増加します。
3. 水の単位体積あたりで計算される一次生産値のレベル(垂直方向のプロファイルに沿った最大値)は、光合成プランクトンの量(クロロフィル含有量)と深さにわたる積分 - 光領域の特性によって決まります。 、貯留層の光合成ゾーンの力を決定します。 その結果、比較的少量の放射エネルギーを吸収する淡い透明な水では、貯水池に入るクロロフィルと太陽エネルギーの単位あたりのプランクトンの一次生産量が増加します。 , 太陽放射の摂取量の増加により、南に向かうにつれて植物プランクトンの一次生産の年間平均値が増加します. 逆に、ツンドラ湖では、光エネルギーの不足によって一次生産が制限されます。地平線より上の低い太陽高度にある貯留層と、氷のない期間が短期間であること。
4. 光合成の光依存性の研究では、水中照射の強度に対する、研究された貯留層に典型的な珪藻と藍藻の群集の反応に類似性があることが示されました。 この関係の分析的表現が見つかり、一次生産量を決定するための計算方法を改善することが可能になりました。
5. ルイビンスク貯水池で得られたクロロフィル含有量に関する長期データは、年々変動を背景にクロロフィル含有量が増加していることを示しており、e-
トロフィーの水溜め。 これは、海岸の工業開発された地域からの栄養負荷の増加と一致しています。 しかし、年々顔料濃度が急激に増加しており、
高気圧型の天気は加速を信じさせる -----
このプロセスでは太陽エネルギーの供給量が増加するため、生体要素とともに光も考慮されます^! 貯水池の富栄養化の要因として。
実施された研究は、植物プランクトンの光合成活性とその一次生産物の形成を確保する上で光が重要な役割を果たしていることを包括的に示しています。 また、貯水池に浸透する太陽放射のエネルギーが、水中光合成のプロセスそのものと同様に、数学的モデリングだけでなく正確な物理的方法を使用して記録できることも重要です。 さらに、浮遊性フィトセノーシスに関しては、生息地との密接な相互作用を伴う比較的単純な構造を特徴とし、モデルは非常にシンプルで自然環境に適しています。 これらすべてにより、水生生態系の生産性の研究における機器および自動測定の可能性が広がります。
文学
1. Aleknn O.A.、Semenov A.D.、スコピンツェフ B.A. 陸水の化学分析ガイド // レニングラード、1973 年。
2. Are F.E.、Tolstyakoo D.N. 太陽放射の水への浸透について // メテオロール。 ヒドロル。 1969 年、N 6。P. 53-64。
3. ベル L.N.、チョーラ S.N.、コルニリエフ V.P. 放射線量を測定する装置(フォトインテグレーター) // Physiol. rzst、1959.T.6、発行。 4. 504-507 ページ。
4. ベレズキン V.A. 光量測定におけるフィルターの応用と、フィルターを使用して観察を処理する方法 // Geophyeich. 1932 年土曜日、T.5、E 号。
5. ブイヨン V.V. 陸水生態系における一次生産のパターン。 サンクトペテルブルク、1994 年。
6. ビリンキナ A.A.、トリフォノバ N.A. 生物起源の要素 // ヴォルガとその生涯。 L.、1978、80-92 ページ。
7. ヴィンバーグ G.G. 湖の水塊における光合成と呼吸を研究した経験。 有機物のバランスの問題について。 メッセージ 1 //Tr. リムノール。 美術。 コシンで。 1934年発行。 18. P.5-24。
8. ヴィンバーグ G.G. 植物プランクトンの量的発達の指標としてのクロロフィル含有量 // 第三の生態学。 会議 報告書の要旨 パート 4。キエフ、1954 年。P. 70-74。
9. ヴィンバーグ G.G. 貯水池の一次生産。 ミンスク、1960年。
10. ヴィンバーグ G.G.、シブコ T.N. プランクトン中のクロロフィル含量の測定 II Izv. BSSR 科学アカデミー。 1953 年、N 3。P. 61-74。
11. ヴィンバーグ G.G.、シブコ T.N.、コバレフスカヤ R.Z. プランクトンのクロロフィル含有量を測定する方法とその適用結果 // 海および内水の一次生産。 ミンスク、1961 年、231-240 ページ。
12. ゲラシモワ N.H. 長期データに基づくサラトフ貯水池とヴォルゴグラード貯水池における植物プランクトン生産の動態 II 科学的 tr. 湖沼研究所 そしてスピーチ。 魚 X-VA。 1981、N 168、P. 65-S1。
13. GOST 17. 1.04.02.-90。 水。 クロロフィル「a」の電気光度測定の方法論 //M.、1990。
14. ダレチナ I.N. 1965年から1967年にかけてのヴォルゴグラード貯水池における有機物の主な生産と破壊。 ⅡTr. サラトフ県 ゴスニオル。 1971.T.10。 47-59ページ。
15. ダレチナ I.N. 1968 年のヴォルゴグラード貯水池における有機物の主な生産と破壊 // ヴォルゴグラード貯水池。 サラトフ、1976 年、55-58 ページ。
16. ダレチナ I.N.、ゲラシモワ N.A. 植物プランクトンとその産物 ヴォルゴグラード貯水池の漁業開発と生物生産の可能性。 サラトフ、1980 年、31-38 ページ。
17.ジュバン A.N. 有機物の主な生成と破壊。 1971 年のサラトフ貯水池の水中の物質。 - // 生物学内部。 水、知らせてください。 速報 1975. N 25. P. 12-15。
18.ジュバン A.N. 1972 年のサラトフ貯水池の水中の有機物の主な生成と破壊。 II 内部の生物学。 水 知らせる。 速報 1976. N 31. P. 13-16。
19. ジュバン A.N. サラトフ貯水池の水中の一次生成、有機物の破壊およびバクテリアの数 // ギドロビオール。 雑誌 1977. T. 13, N 1. P. 81-85。
20. ダヴギー T.N. バイカル湖の水中太陽放射。 ノボシビルスク、1977年。
21. エヴネビッチ.T.V. 大気の透明度特性の計算について // メテオロール。 ヒドロル。 1984. N 5. P. 103-109。
22.エリザロワV.A. ルイビンスク貯水池 II ハイドロビオールの水中の植物色素の組成と含有量。 雑誌 1973.T.9、N2。 23-33ページ。
23. エリザロワ V.A. ルイビンスク貯水池の植物プランクトンバイオマス単位当たりの光合成色素の含有量 // ヴォルガ川の動植物および微生物。 ルイビンスク、1974 年、46-66 ページ。
24.エリザロワV.A. 1970年の観察によるイワンコヴォ貯水池の植物プランクトン色素の含有量 // 水生生物の生物学、形態学および系統学。 L.、1976。P. 82-90。
25.エリザロワV.A. ルイビンスク貯水池における植物プランクトン色素の季節動態と分布 // 下等生物の生物学と系統学。 L.、1978.S. 103-124。
26. エリザロワ V.A.、シガレワ L.E. ルイビンスク貯水池の浅海における植物プランクトン色素の含有量 II 上部アオルガ貯水池の沿岸浅海の水文学的状況。 ヤロスライル、1976 年、133 ~ 147 ページ、
27. ザイドル A.M. 物理量の測定における誤差。 L.、1974年。
23.イヴァトシュ」A.B. 1965 年頃のクイビシェフ貯水池の微生物学的特徴。および微生物学、1968。T.37、no. 2. 360-366 ページ。
29. イヴァチン A.B. 「8ob g.// biolgich gnutr.」sod におけるクイビシェツク医療システムにおける有機物の主要な生産と破壊. Inform. s>yul. 1970. M 8, pp. 15-19.
30. イヴァチン A.B. クイビシェフ貯水池における植物プランクトンの生成と有機物の破壊 // ギドロビオール。 雑誌 1974. T.10、N 3. P. 65-69。
31. イヴァチン A.B. 有機物の生成と破壊、およびクイビシェフ貯水池。 L.、1983。P. 90-102。
32.キリロワTV 湖と貯水池の放射線体制。 L.、1970年。
33. コブレンツ・ミシュケ O.I. 種子の吸収と廃棄
黒海の植物プランクトンの光合成のプロセス N 黒海の生態系。 M.、1E30。 132~139ページ。
34. Ksblznts-Uishye O.I.、Voznik V.、Ochakovsky Yu.E. (編)。 /p&osik* 太陽エネルギー z プロセス* foshsknteza chgomomsrsk%:g.. ■l -¿Altai fpuplaneten。 M.、IS85。
25. 「.Soblenz-Mishkp O.I.、Vogiyak B.、Konooanoye O.V. Md“-P17sg.;;”すなわち..r&sy-」 -gchnch^och products // Oi.vawrewm Baptik"! 。 >『非イスナ1984年。 N.、.037。 pp.385-40/
l. ラリン D.A. üiyöb ßQAOSiAoa.7 üöcihhk MSU に侵入する Sopntsp の死亡計算について。 シリーズ V. 地理。 1973. N 1. S. 102-106。
37. ムスウェフト「H.f」1. Perps.chnpya プラットロン FWHihghov g.ndohrlnchlishcha // サポートと croduttnod pzd&gich&mmkh およびヴォルガ盆地貯水池の沿岸植物虫害の生産。 L.、1990。S. 207-216。
38. モキアスキー K.A. ラドガ湖のヨウ素塊への太陽の放射エネルギーの浸透 // ラドガ湖の熱オヤシム。 L.、1968。P. 73-99。
39. モキフスキー K.A. オネガ湖の放射線バランス II オネガ湖の熱体制。 L、1973。P. 52-134。
40. ニチロロヴィッチ A.A. 光合成と高収量を得る理論。 XVティミリャゼフの読書。 M.、1956 年。
41. ニチロロヴィッチ A.A. 生物圏および農業における生産性の基礎としての植物の光合成活動 // 光合成と生産プロセス。 M.、1988。P. 5-28。
42. ポカチロバ T.N. 自然水域における太陽放射のスペクトル減衰 II バイカル湖およびその他の水域の水文学。 ノボシビルスク、1984年。14-19ページ。
43. ポカチロバ T.N. 日射計を使用して水中太陽放射のエネルギーを測定する // 内陸水域におけるプランクトンの一次生産を研究する際の方法論的問題。 サンクトペテルブルク、1993 年、139-146 ページ。
44. プリマチェンコ AD 植物プランクトンの光合成生産と川の栄養状態。 エニセイ // エニセイの生産的および水生物学的研究。 ノボシビルスク、1993 年、149-161 ページ。
45. Razgulin S.M.、Gapeeva M.V.、Litvinov A.S. 1980 年のルイビンスク貯水池の栄養素とイオンのバランス。 II ヴォルガ貯水池の水化学研究。 ルイビンスク、1982 年、61-91 ページ。
46.ローゼンバーグ。 G.V. 光散乱物質の分光法の物理的基礎 // 物理学、科学の進歩。 1967. T.91、N 4. P. 569608。
47. ロマネンコ V.I. ルイビンスク貯水池における有機物の形成と破壊の微生物学的プロセスの特徴 II 内陸貯水池における有機物の生産と循環。 M.-L.、1966 年。P. 133153。
48. ロマネンコ V.I. ルイビンスク貯水池における植物プランクトンによる有機物の生成 // ジドロビオール。 雑誌 1971.T.7、N4。 5-10ページ。
49. ロマネンコ V.I. 内陸水域における有機物の生成と破壊の微生物学的プロセス。 L.、1985年。
50.ルトコフスカヤ 8.A. 湖、貯水池、海の放射線特性の特徴 I/海洋気象研究。 M..19S2。 117-132ページ。
51. ルトコフスカヤ V.A. 陸と海の水域への塩分放射線の浸透 II 太平洋とインド洋の大気循環と表面空気層の研究。 M.、1965。S. 245-274。
52. サルマノフ M.A.、ソローキン Yu.I. クイビシェフ貯水池の一次生産 // Izv。 ソ連科学アカデミー、シリアル。 バイオル。 1962. N 4. P. 603-613。
53.サッポL.M. 1974年から1976年にかけてイワンコヴォ貯水池で有機物の主な生産が行われました。 // 土曜日 科学的 tr. 湖沼研究所 そしてスピーチ 魚 X-VA。 1981. N 105. 30-34 ページ。
54. Semenchenko!?.A. (Meleshko I.E.、Kirmalov R.L.、Vladimirova I.A. リモコン付き水中記録分光計 // Meteorol. Hydol. 1971. N 4. P. 9499.
55.シガルヴァ L.E. ゾルガ上部の植物プランクトンのクロロフィル含有量と光合成活性。 ディスる。 博士号 バイオル。 科学。 ボロック、1984年。
56. ソローキン Yu.I. ルイビンスク貯水池の水柱における有機物の主な生成 // Tr。 バイオル。 美術。 「ボロック」 1958. Vol. 3. 66-68 ページ、
57. Sorokin Yu.I.、Rozanova E.P.、Sokolova G.A. Sm //Tr を使用したゴーリキー貯水池の一次生産の研究。 全て ハイドロビオール。 約-va。 1959.T.9。 351-359ページ。
58. ソローキン Yu.I.、フェドロフ V.K. オネガ湖における有機物の主な生産と破壊の決定 // 水生生物の生理学と有機物の循環におけるそれらの役割。 L.、1969。P. 4-8。
59. ソ連の気候に関するハンドブック。 L.、1966。Vyl.8、パート 1。 問題 12、パート1。 第 13 号、パート 1。 第 29 号、4.1。
60.ストルブノワ V.N. 動物プランクトン // プレシチェエヴォ湖の生態系。 L.、1989、156-173 ページ。
61. タラセンコ L.V. 70年代のイワンコヴォ貯水池における植物プランクトンの生産特性。 M.、1963 年。 (原稿は 1983 年 6 月 8 日に VINITI に寄託 N 3153-83 寄託)
62. タラソワ T:N. 河川における有機物の主な生成と破壊。 1956 年のチェボクサル水力発電所の建設ルート上のヴォルガ。」 // Uch. ザップ。 ゴルコフスク。 ウンタ。 サー。 バイオル。 1970年発行。 105. 32-36 ページ。
63. タラソワ T.N. ゴーリキー貯水池とゲドロビオールにおける一次生産、バクテリプランクトン、有機物の破壊。 裁判官 1973. T.9、N 3. P. 5-11。
64. タラソワ T.N. ゴーリキー貯水池の湖部分における有機物の破壊における細菌の一次生産と関与 // 貯水池における物質とエネルギーの循環。 生物要素 循環。 バイカル湖のカラマツ。 1977 年、69-73 ページ。
65.チェキンLP 貯水池の明るい体制。 ペトロザヴォーツク。 1987年。
66. Shmelev G.A.、Subbotina E.V. 1974 年から 1979 年のゴーリキー貯水池における光合成による有機物の生成と破壊。 // 土曜日 作品 Gork.、Volzhsk、および Rybin、ヒドロメテオロール。 観察者。 1983. N 16. P. 51-59。
67.ヤニシェフスキーYu.D. 光濃度測定装置とそれを用いた観察方法。 L.、1957年。
68. バニスターT.T. リムノイの適応を含む、定常状態、栄養飽和状態の藻類の成長の定量的説明。 海洋学者。
1979年。 24、N 1。P. 76-96。
69. バークホルダー P.R.、バークホルダー L.M.、リベラ J.A. 一部のサンゴや海洋植物に含まれるクロロフィル「a」 // 自然。 1959年。 183、N 4671。P. 1338-1339。 > "
70. Curtis J.T.、Juday C. ウィスコンシン州の湖における藻類の光合成 // インターン。 牧師 ゲス。 ハイドロビオイ。 1937 年。 35、H.2。 S. 122-133。
71. ガンフ G.G. 赤道の浅い湖(ウガンダ、ジョージ湖)からのファイトピンクトンの光合成生産と放射光と光合成の関係 // Oecologia。 1975年。 18、N 3。P. 165-183。
72. Ganf G.G.、Heaney S.I.、Cony J. 非気体空胞性シアノバクテリア Oscillatoria bourretlyl (Tychoma bourretlyl) の自然集団および培養物における光吸収と色素含有量、および J. Plankton Res. 1991.Vol. 13、N5。 P.1101-1121。
73. ゲスナー F. ハイドロボタニック。 Bd.1。 ベルリン、1955 年。
74. ヒーニー S.4.、テーリング J.F. Ceratlum hyrundlnelta - 複雑で移動可能で成功を収めた植物の生態 II Rep. 淡水バイオル。 准教授
1980. N 48. P. 27-40。
75. ハンフリー GF 太平洋におけるファイトピンクトン色素 II Proc. 会議 海洋および淡水の一次生産性測定に関する講演が大学で開催されました。 ハワイ、1961年8月21日~9月6日。 私たち。 原子力委員会、1963 年。p. 121-141。
76. 市村 S 有賀 Y. 淡水植物ピンクトンの光合成のいくつかの特徴 // Bot. 東京、マグニチュード。 1958. Vol. 71、N 841-842。 P.261-269。
77. ジェフリー S.W.、ハンフリー G.F. 高等植物、藻類および天然植物ピンクトンのクロロフィル「a」、「b」、「ci」および「сг」を決定するための新しい分光測光方程式 II Biochem. 生理。 フィアンツ。 1975。BD。 167、N2。 P.191-194。
78. ジュウソン D.H. よく混合された湖(北アイルランド、ネイ湖)における正味の光合成を制御する成分の相互作用 II Freshwater Biol。 1976年。 6、N 6、P.551-576。
79. ジュウソン D.H. アイルランド北部、ネイ湖の有光地帯//オイコスの植物ピンクトン含有量に関連した光の透過。 1977年。 28.N 1.P. 74-83。
80. ジューソン D.H.、テイリング J.F.、ドリング M.J.、ティルザー M.M.、ヒーニー S.I.、カニンガム C. 淡水中の光合成的に利用可能な放射線の測定: 一次生産の研究に使用されるいくつかの現在の機器の比較テスト II J. Plankton Res. 1984. Vol .6、No.2、P.259273。
81、カーク J.T.O. 水生生態系における光と光合成。 ケンブリッジ。 1983年。
82. コスキ V.M.、スミス J.H.C. 黄化した大麦苗からのプロトクロロフィルの単離とスペクトル吸収特性 II J. Am. 化学。 1948年。 70. P. 3558-3562。 _________________
83. ローレンゼン C.J. クロロフィルと褐色色素の測定: 分光測光方程式 // Llmnol. 海洋学者。 1967年。 12、N 2。P. 343-346。
84. ロロンゼン C.J.、ジェフリー S.W. 海水中のクロロフィルの測定 II 海洋科学におけるユネスコ技術文書。 1980、N 35 P. 3-20。
85. マニング W.M.、ジュデイ R.E. ウィスコンシン州北東部のいくつかの湖のクロロフィル含有量と生産性 // Trans。 賢い。 アカド。 科学。 1941年。 33. P. 383-392。
88. Marfwr A.F.H.、Kusch E.A.、Ra! H.、Rfsmann 3. 淡水における合成シルク色素の蓄積と方法の標準化: 結論と推奨事項 // Arch. Hydroblol.、Beih.Ergebn. イルムホル。 1980. H. 14. P. 91-106。
87. モレス C.M.、イェンチュ C.S. 海洋植物プランクトンからのクロロフィルの分解におけるクロロフィラーゼと光の役割 H J. Ex>。 3月 バイオル。 エコル。 1970年。 4、N 3、P.238-249。
88. ニール P.J.M. テイリング J.F.、ヒーニー S.I.、レイノルズ C.S.、ルンド J.W.6。 イギリス湖水地方からの長期時系列: 春の最大期における日射量依存性の植物プランクトンの動態 // Llmncl. セノグル。 1991.Vol. 36、N 4、P.751-760。
09. パーソンズ T.R.、ストリックランド法務博士 クロロフィルとカロテノイドを確認するための改訂された方程式を用いた海洋植物色素の分光光度測定に関する議論//J. 3月 解像度 1983.Vol. 21、N3。 P.155-163。
それで。 Plait T.、Galiegos C.L.、Harrison W.G. 海洋植物プランクトンの自然集団における光合成の光阻害 // J. Mar. 解像度 1980. Vol. 38、N 4。P. 687-701。
91. Piatt T.、Sathyendranath S.、Ravindran P. 植物プランクトンの一次生産: 水面の単位面積あたりの日量の分析ソリューション//Proc. R. Soc. ロンドン。 1990.Vol. 241. P. 101-111。
92. 光合成放射エネルギー II 海洋科学におけるユネスコの技術文書に関するこの合同専門家グループの第 1 回会合の報告。 1965.N 2.
93. リチャーズ F.A.、トンプソン TG. 色素分析による植物プランクトン個体群の推定と特性評価。 ft.プランクトン色素の推定のための分光測光法 //J. 3月 解像度 1952. Vol. 11、N 2、P.156-172。
94. ショーマー H. ウィスコンシン州北東部の湖のさまざまな深さにおける水生植物の光合成 // 生態学。 1934年。 15、N 2、P. 217218。
95. Schomer H.A.、Juday C. ウィスコンシン州北東部のいくつかの湖のさまざまな深さでの藻類の光合成。 I. 1933 年の観察//Trans。 賢い。 アカド。 科学。 1935年。第29巻。 P.173-193。
96. SCOR-ユネスコ作業部会 N 17. 光合成色素の測定 II 海水中の光合成色素の測定。 海洋学の方法論に関するモノグラフ。 1966.1。 P.9-18。
97. Stra^krafaa M.、Gnauk A. 淡水生態系 - モデリングとシミュレーション。 アムステルダム。 1985年。
98. スミエック M. 池に浸透する太陽放射に対する制御された富栄養化の影響 II Acta Hydroblol。 1975年。 17、N 2. P. 149-182.(no/ibCK.)
99. J.Fを尾行する 一部の淡水珪藻の光合成特性 水中放射線との関連 II New Phytol. 1957年。 56、N. 1. P. 29-50。
100. J.F.の尾行 自己シェーディング効果 プランクトンの自然個体群における珪藻 // より濡れやすくなります。 レーベン。 1960年。ジャーグ。 12、H.9-10。 S. 235242。
101. Tailing J.F.、淡水植物ピンクトンの生産生態における制御因子としての水中光気候 // Mitt、int。 ヴェレイン、テオール。 アンギュー。 イルムノル。 1971、N 19、P. 214-243。
102. J.F.の尾行 ファイトピンクトンによる太陽放射の利用 // 光生物学の動向、ニューヨーク、1982 年、P. 619-631。
103. J.F.の尾行 英国の 4 つの湖流域における植物ピンクトン総バイオマスの 26 年間の記録における季節変化と年ごとの変動性と安定性の比較 // Hydrobiologla。 1993.Vol. 268、N1。 P.65-98。
104. Tailing J.F.、ドライバー D. 植物ピンクトンのクロロフィル "a" の推定におけるいくつかの問題 // Proc、Conf。 海洋および淡水の一次生産性測定に関する講演が大学で開催されました。 ハワイ、1961年8月21日~9月6日。 米国原子力委員会。 1963 年。P. 142-146。
105. テーリング J.F.、ウッド R.B. プロッサー M.V.、バクスター R.M. ファイトピンクトンによる光合成生産性の上限:エチオピアのソーダ湖からの証拠 // 淡水バイオール。 1973年。 3、N 1. P. 53-76。
106. トゥルザー M.M. ボーデン湖 II リムノールにおける植物ピンクトンの生産性に対する光合成色素による部分光吸収の重要性。 海洋学者。 1983.Vol. 28、N 5。P. 833-846。
107. ティルザー M.M. 植物ピンクトン // Arch の垂直光合成プロファイルを制御する基本的なパラメーターとしての量子収量。 ハイドロバイオール、サプリメント。 1984 年。 69、N 2. S. 169-198。
108. Tilzer M.M.、Schwarz K. 高山の湖における植物プランクトンの光適応の季節的および垂直的パターン // Arch。 ハイドロブロール。 1976.BD。 77、N 4.5.488-504。
109. トム・アッソン K. 北極と高山の湖に関する考察 II オイコス。 1958. Vol. 7、N 1. P. 117-143。 ____________________________
110. フォーレンヴィーダー R.A.、テーリング J.F.、ウェストレイク D.F. 水生環境における一次生産量の測定方法に関するマニュアル。 オックスフォード。 1974年。
111. イェンチュ C.S. 藻類培養物中のクロロフィルを定量的に推定するための非抽出法 // Nature。 1957>Vol. 179、N
4573. P. 1302-1304。
112. イェンチュ C.S. 葉緑素顔料の分解生成物H Memの測定を参考にしたクロロフィルと光合成炭素生成の関係 1位。 イタル。 イドロビオール。 補足 .13., 1255. P. 323-3「インチ」
学位論文のテーマに関する主な出版物
ピリナ I.L. 季節の照明と関連した藻類の光合成の強さ // Tr. 研究所 バイオル。 貯水池 1959.T.1(4)。 102-109ページ。
2.ピリナI.L. ヴォルガ川とその貯水池の光合成産物 // 速報。 イクスト。 バイオプ。 貯水池 1959年(a)。 N 3. 17-20 ページ。
3. ピルンズ I.L. 植物プランクトンの組成に対する一次生産の依存性 // 海と内水の一次生産。 ミンスク、1961 年。P. 30.8-313。
4.ピリナI.L. 暫定結果、淡水植物プランクトンの色素を決定するための分光測光法の応用 // 貯留層研究の生物学的側面。 M.-L.、1963 年。P. 51-59。
5.ピリナ I.L. 貯水池内の異なる光条件下での淡水植物プランクトンの光合成 // レポートの要約。 1回の会議 湖の貯水池における物質とエネルギーの循環の問題について。 バイカル湖のリスティーンシャ、1964 年。P. 81-62。
6.ピリナI.L. 水中フォトインテグレーター // ギドロビオール。 雑誌 1365、T.1、N 2. P. 61-07、
7.ピリナI.L. クロロフィル含有量と水中放射線の強度の測定結果に基づく植物プランクトンの一次生産の評価 // 水生物学の問題 (全ロシア国立植物協会の第 1 回会議の要約) Mn 1965 (a)。 359-360ページ。
8. Pyrina I.L.、Feoktistova O.I.、貯水池内の有機物の生産における藍藻の役割 // 藍藻の生態と生理学。 M.-L.、1965 年。P. 42-46。
9. ピリナ I.L. いくつかの要因に応じたイワンコボ、ルイビンスク、クイビシェフ貯水池における植物プランクトンの一次生産 // 内陸貯水池における有機物の生産と循環 - L.、1966。P. 249-270。
10.ピリナ I.L. 照明、クロロフィルおよび植物プランクトンバイオマスに関連したいくつかの貯留層の一次生産 // Ph.D. の要約 ディスる。 ミンスク、1966年(a)。 16ページ
11. ピリナ、イリノイ州、マケエバ、N.L. メンブランフィルター上で収集された藻類細胞のクロロフィルを測定する方法について // ヴォルガ貯水池の植生。 M.-L.、1966年。P. 198202。
12. イリノイ州ピリナ。 貯水池におけるさまざまな光条件下での淡水植物プランクトンの光合成 // 湖の貯水池における物質とエネルギーの循環。 M.、1967。S. 202-208。
13. ピリナ I.L. ヴォルガ貯水池におけるプランクトンの光合成中の太陽エネルギー利用効率 // 水生生物の生活における放射因子。 L.、1967 (a)。 34-42ページ。
14.ピリナ I.L. 植物プランクトンの光合成のバイオマスおよびクロロフィル含有量への依存性 // 内陸水域の微生物相、植物プランクトンおよび高等植生。 L.、1967 (b)。 94-103ページ。
15.ピリナ I.L. ヴォルガ上流および中流の貯水池における一次生産 // レポートの要約。 私は確信します。 ヴォルガ盆地の貯水池の研究について。 トリヤッティ、1968 年、88-80 ページ。
16.ピリナ I.L. 光学的に異なる水域における太陽光の強度とスペクトル組成 // 第 2 回会議。 湖の貯水池における物質とエネルギーの循環の問題について。 報告書の概要、パート I。Listvenichnoe on Baikal、1969。83-84 ページ。
17. ピリナ I.L.、エリザロワ V.A. いくつかの淡水域の植物プランクトンのクロロフィル含有量 // 第 2 回会議。 湖の貯水池における物質とエネルギーの循環の問題について。 レポートの概要、パート II。 バイカル湖のカラマツ、1969 年。9-10 ページ。
18.ピリナ I.L. 貯水池の一次生産の指標としての水中照明とクロロフィル // 海洋および大陸の貯水池における生物学的プロセス。 (VSBO の報告書と会議の論文)。 キシナウ、1970 年、P. 314。
19. ピリナ I.L.、エリザロワ V.A. 一部の藻類の培養物中のクロロフィルの分光光度測定 II 淡水生物の生物学と生産性。 L.、1971、P. 5665。
20. ピリナ I.L.、ナウモバ E.I. コナコヴォ州地区発電所の廃水における植物プランクトンの光合成活動 // Tezi-
シドクル。 症状 火力発電所の水が水域の水文学と生物学に及ぼす影響について。 ボロック、1971年。 P.52
21. Guseva K.A.、Priymachenko A.D.、Pyrina I.L. Kozhova O.M.、Gerasimova N.A.、Kuksn M.S.、Shalar V.M. ソ連の水力発電所の貯水池における藍藻の発生 // 報告書の要約。 全て ミーティング ため池の総合利用について。 キエフ、1971 年。P. 41-42。
22. ピリナ P.L.、ルツクスフスカヤ V.A.、イリンスキー A.L. ヴォルガ貯水池の水への太陽放射の浸透に対する植物プランクトンの影響について II ヴォルガ貯水池の水文体制の有機物と要素。 L.、1972。S. 97-106。
23. ピリナ I.L.、ゲッセン M.V.、エリザロワ V.A. ツンドラのハルベイ湖における植物プランクトンの生産性のいくつかの指標 // 湖と貯水池における物質とエネルギーの循環 (第 3 回 Soyaeshchanke.. 報告書の概要、コレクション 1)。 バイカル湖のリストヴェニクノア、1973 年、112-114 ページ。
24. ピリーナ Sh1.、エリザロワ V.A.、ニコラエフ I.I. オネガ湖の植物プランクトン中の光合成色素の含有量と、この貯水池の生産性レベルを評価するためのそれらの重要性 II オネガ湖の生産性。 L.、1973。S. 108-122。
25. ピリナ I.L.、ナウモバ E.I. コナコボ州地区発電所の廃水における植物プランクトンの光合成の強さ // 速報。 研究所 バイオル。 内部 水 M に知らせる 速報 1973. N 17. 18-22 ページ。
26. Pyryaka Ya. L. 光フィルターを備えたフォトインテグレーターを使用した光合成波長範囲における水中放射線のスペクトル測定 II 大気および地表における放射線プロセス (光量測定に関する IX 全連合会議の資料). L., 197-1. P.423-428。
27.ピリナ I.L. ヴォルガ貯水池における植物プランクトンの一次生産 // レポートの要約。 II 会議 ヴォルガ流域の貯水池の研究に関する(報告書の概要)。 ボロック、1974年(a)。 20-24ページ。
28. ピリナ I.L. 植物プランクトンの一次生産 (モノグラフのセクション) II 内陸水域の生物地球変動を研究する方法。 M.、1975。S. 91-107。
29. ピリナ I.L. 光学的に異なる水域における太陽光の透過とスペクトル組成、および湖の貯水池における物質とエネルギーの循環。 ノボシビルスク、1^75 (a)。 349-353ページ。
ZO^Pyrika I.L.、Elizarova V.A. いくつかの淡水域の植物プランクトンのクロロフィル含有量 II 湖沼貯水池における物質とエネルギーの循環。 ノボシビルスク、1975年。P. 85-89。
31. ピリナ I.L.、エリザロワ V.A. さまざまな種類の貯水池における光合成の強度と塊状の植物プランクトン中のクロロフィルの含有量 // 水資源の研究と保護。 M.、1975 (a)。 P.60。
32. ピリーナ I.L.、デビヤトキン V.G.、エリザロワ V.A. 植物プランクトンの発生と光合成に対する加熱の影響に関する実験的研究 // 貯水池の寿命における人為的要因。 L.、1975。S. 67-84。 、
33. ピリナ I.L.、バシュカトワ E.L.、シガレバ L.E. 1971年から1972年にかけてのルイビンスク貯水池の浅いゾーンにおける植物プランクトンの一次生産 // ヴォルガ上流貯水池の沿岸浅瀬の水文学的状況。 ヤロスラヴリ、1976 年、106-132 ページ。
34. ピリナ I.L.、ゲッセン M.V.、ヴァインシュタイン M.B. ボリシェゼメルスカヤ ツンドラのハルベイ水系の湖における植物プランクトンの一次生産(モノグラフのセクション) // ボリシェゼメルスカヤ ツンドラ東部の湖の生産性。 L.、1976。P. 63-76。
35. Elizarova V.A.、Pyrina I.L.、Getsen M.V. カーベイ湖の水中の植物プランクトン色素の含有量。//同上。 P.5563。
36. ピリナ I.L.、ルトコフスカヤ V.A. 植物プランクトンの光合成強度の、水に浸透する総日射量への依存性 // ヴォルガ流域における生物生産プロセス。 L.、1976。P. 48-66。
37. Guseva K.A.、Priymachenko-Shevchenko A.D.、Kozhova O.M.、Pyrina I.L.、Kuksn M.S.、Shalar V.M.、Chaikovskaya T.S.、Gerasimova N.A. ソ連の水力発電所の貯水池における「ブルーム」の強さと藍藻類の大量植生のタイミング // 国民経済における水の「ブルーム」と藻類の使用を規制する問題。 キエフ、1976 年、6-19 ページ。
38. ピリナ I.L.、エリザロワ V.A.、シガレバ L.E. 植物プランクトンの生産性に関するイワンコヴォ貯水池の富栄養化の兆候 I. 天然水の人為的富栄養化。 チェルノゴロフカ、1977 年、238-244 ページ。
39.ピリナ I.L. 植物プランクトンの一次生産(モノグラフのセクション) // イワンコヴォ貯水池とその生活。 L.、1978.S. 102-124。
40.ピリナ I.L. 植物プランクトンの一次生産 (モノグラフのセクション) // ヴォルガとその生活。 L.、1978 (a)。 P.148~152。
41. ピリナ I.L.、バシュカトワ E.L.、ミネバ N.M.、シガレバ L.E. シェクスニンスキー貯水池と隣接する湖における植物プランクトンの生産性に関するいくつかの指標 II ヴォログダ地域の水域の生物資源、その保護と合理的利用
募集(科学と実践の会議の抄録)。 ヴォログダ、1978 年、20-21 ページ。
42. ピリナ I.L.、シガレバ L.E. 1973~1974 年のイワンコズ貯水池の植物プランクトン色素の含有量 // 下等生物の生物学。 ルイビンスク、1978 年、3-17 ページ、
43. ピリナ I.L. ヴォルガ川における植物プランクトンの一次生産 (モノグラフセクション) //ヴォルガ川とその生涯。 ハーグ、1979 年。P. 180-194。
44.ピリナ I.L. 最大光合成、総日射量、水の透明度 // ギドロビオールに基づく植物プランクトンの一次生産量の決定。 雑誌 1979。T. 15、N 6。P. 109-113。
45. ピリナ I.L.、トリフォノバ I.S. ラドガ湖//ギドロビオールにおける植物プランクトンの生産性に関する研究。 雑誌 1979. T. 15、N 4. P. 26-31。
46.ピリナI.L.、エリザロワV.A. クロロフィル含有量に関するさまざまな栄養レベルの搾乳の特性の比較 // 物質循環と水域の生物学的自己浄化。 キエフ、1980 年。P. 100-109。
47. ティルザー M.M.、ピリナ I.L.、ウェストレイク D.F. 植物プランクトンの光合成 (モノグラフのセクション) // 淡水生態系の機能。 ケンブリッジ、1980 年。163-170、174-176 ページ。
48. ピリナ I.L.、ミネバ N.M.、コルネバ L.G.、レタンスカヤ G.I. 植物プランクトンとその生成物 (単行本のセクション) // ソ連北西部の大きな湖に対する人為的影響。 パート II。 水文学
ホワイト湖の水素化学。 P.、1981。P. 15-64。
49. ピリナ I.L. 氷下の植物プランクトン植生の光下限界について // バイカル湖地域の生態系の問題!!。 水域の細菌数の監視。 イルクーツク、1982 年、85-86 ページ。
50. Pyrina I.L.、Smetanin MM: 植物プランクトンの量的特徴の貯留層の平均値の評価について /■ 「水生生物学および魚類学的研究における誤差の推定。Rybinsk、1982、pp. 144-156」。
51..ピリナ ^ I.L. 。 氷の下を透過する光合成活性放射線の研究 //放射線気候学と
光量測定の応用側面 (光量測定に関する XI! 会議の資料)。 イルクーツク、1984 年、252-254 ページ。
52、ピリナ I.L. エオルガ盆地上部の貯水池における可視日射量と総日射量との関係//貯水池内の物質とエネルギーの循環。 イルクーツク、1985 年、38-40 ページ。
53.ピリナ I.L. 北西部の大きな湖貯水池における氷河期の光体制の条件と植物プランクトンの発達 // ソ連の大湖の研究の問題。 L 1985 (a)。 111-114ページ。
54. ピリナ I.L.、シガレバ L.E. さまざまな水文気象条件におけるルイビンスク貯水池の植物プランクトン色素の含有量(1972~1976年) // 水生生物の生物学と生態。 L.、1986。P. 65-89。
55. ミネバ N.M.、ピリナ I.L. ルイビンスク貯水池における植物プランクトン色素の研究および同上。 90-104ページ。
56. ピリナ I.L.、ミネバ N.M.、シガレバ L.E. ルイビンスク貯水池における植物プランクトン色素の長期研究 // V VSBO 会議、パート I。報告書の要約。 クイビシェフ、1986 年、208-209 ページ。
57. プリマチェンコ AD、ピリナ I.L.、ヴォチャコワ N.E. 川のプランクトンと底生生物の色素組成と光合成活動。 エニセイ // 同上、パート II。 254-285ページ。
58. ピリナ I.L.、ミネバ N.M.、シガレバ L.E.、バロノフ I.M.、マジン A.M. プレシチェエヴォ湖の植物プランクトンの生産性について // ロシア平原の中央部と北部の内水の保護と合理的利用。 ヤロスラヴリ。 1986。36-40 ページ。
59.ピリナ I.L. 自然水域における太陽放射の弱まりについて II 第 1 回ヴェレシチャーギン・バイカル国際会議。 レポートの要約。 イルクーツク、1989 年、35-36 ページ。
60.ピリナ I.L. 自然水中での日射量の減衰について。 "//。第一回ベレシャギン・バイカル国際会議。要約。イルクーツク、1989、P. 35-36。
61.ピリナ I.L. 水力光学特性と光体制 (モノグラフのセクション) II プレシチェエヴォ湖の生態系。 L.、1989 (a)。 S.3540」
62. ピリナ I.L.、シガレバ L.E.、バロノフ I.M. 植物プランクトンとその生産能力 // 同上。 71-114ページ。
63. ピリナ I.L.、シガレバ L.E.、ジュバン A.N. 植物プランクトンの一次生産 // 同上、114-122 ページ。
64. ビクブラトフ。 E.S.、ジュバン A.N.、ピリナ I.L. 有機物のバランス//同上、216-218 ページ。
65. ジュバン A.N.、ピリナ I.L. 栄養状態と水質 // 同上。 218-221ページ。
66. ピリナ I.L.、ミネバ N.M. ルイビンスク貯水池//植物相の水柱中の植物プランクトン色素の含有量と、ヴォルガ盆地の水域の遠洋および沿岸植物の生産性。 L.、1990.S. 176-188。
67.ピリナ I.L. ルイビンスク貯水池におけるクロロフィルと植物プランクトンの生産性の長期動態 // 植物の成長と生産性の調節の生態学的側面。 ヤロスラヴリ。 1991 年、253-259 ページ。
68.ピリナ I.L. 湖内の植物プランクトンのクロロフィル含有量と生産性。 プレシチェエヴォ // プレシチェエヴォ湖の富栄養化の要因とプロセス。 ヤロスラブリ、1992年、18-21ページ)。
69. ピリナ I.L.、リャシチェンコ O.A. 湖における植物プランクトンの組成と生産性。 現段階のプレシチェエヴォ。 // レポートの要約。 全ロシア人 科学的 国内労働組合300周年を記念した会議。 Pereslzl-Zalegky、1992 年、112-114 ページ。
70. ピリナ I.P.、リャシェンコ O.A. 湖の植物惑星の構成と生産性。 現段階のプレシチェフ。 // トリさん。 全ロシア人 泉さん、構成。 ロシア艦隊創設 300 周年に捧げられました。 yyl.Sh. シュレスラヴリ・ザルスキー。 1392。48-54 ページ。
71.イリノイ州ピリナ。 nepciv-mcfi 植物プランクトン生産を決定するための酸素法 // 内部水域におけるプランクトンの一次生産を研究する際の方法論的問題。 Syacht-Petersburg、1053、10-13 ページ。
72 プムリナ I.L. 光合成の条件と分離サンプルの特徴; 植物プランクトン。 // 同上。 21-23ページ。
73.ピリナPL。 水中光合成反応の測定
1IPM11L MYAP1LYAIMM N LOII WAf
74. Mbtjjwha i"i.."!.、vf.-.eiaiiiiHM.fvi。 一次生産量の積分値と平均値の計算とその精度の評価。 // 同上。 30~40ページ。
75. イリノイ州ピリナ。 ヴォルガ植物プランクトンの一次生産に関するデータの遡及的分析 // 大河川流域の生態学的問題。 トリアッティ、1993 (a): 129-130 ページ。
78. ピリナ I.L.、プリマチェンコ AD 川中の植物プラネトン色素の含有量。 エニセイ (モノグラフのセクション) // エニセイの生産的および水生物学的研究。 ノボシビルスク、1993年。P.
7-" Perina I..P.、mineeza N.M.、Cnrfepfetiä l.E,. .imnao;: ; A D Filimonov V.S.、Mitropolskal M.U. 空間的に? .."spso-children's phytoplankton (p^zdol." :> 生態学者 rfi-wi, ~ FOGT.5rY prog.trlysteennogo RYSPRODSG.SIIK L.trenvshcheima (ID-roOkontov St. Petersburg, 19EZ. P. 55-8-1. 70. Pyrina I.L., Smetaniya M.M. ., Smetznnnna TL. Sgatshshmvsk)」濃度を評価するアプローチ植物プランクトン遺伝子の色素とバイオマスの評価および植物プランクトン生産性の評価、Noeosibirsk、1993、P. 30-4 4.
79.ピリナ I.L. 植物プランクトンの一次生産の指標
ケシュデ貯水池川。 ヴォルガ // 内陸水域における微生物叢と生物学的浸透」。サンクトペテルブルク(印刷物について)。60。Pyrina I.L.、Dobrynin E.G.、Mineeva N.M.、Sigareva L.E. 成層の酸素体制に対する光合成と有機物の破壊の影響についてlake. Ibid. 81. Pmpiche I.L., M^meeva N.M. ルイビンスク貯水池の太陽光の浸透と光合成ゾーンのパラメーター // ルイビンスク貯水池の植物プランクトンの構造と生産性. Tolyatti (印刷中)-
植物プランクトンの生産サイクル
植物プランクトンの一次生産を決定する主な要因は、光層の厚さ、植物プランクトンへの栄養素の供給、および生産サイクルの期間です。
一般に、これらのパラメータは各地域の特定の気候および水文条件に依存し、痕跡領域と生産サイクルの終わりに密接に関係する春の発達の開始のタイミングに現れます。 さらに、これらのパラメータは、春の発達の初めに光層に形成される栄養素の供給によって決定され、川の流出と冬の深さの対流混合、夏の垂直乱流拡散、太陽エネルギーの量によって決まります(順番に依存します)。季節や気候要因など)、植物プランクトンの構造的および機能的特徴。
白海の植物プランクトンは、「沿岸」着生型と淡水型が大量に混合したバレンツ海の定性的に枯渇したプランクトンです(Kokin、Koltsova、1972)。 白海では、バレンツ海南部と比較して、種数は寒さを好む珪藻が大半を占め、ペリディニア類の種の多様性は減少しています。 白海の植物プランクトン組成に含まれる藻類の裾野(63%)は、カラ海(67%)やラプテフ海(64%)などの高北極の大陸外海に近く、このことも北極の性質を示している。 さまざまな生物学的季節における植物プランクトンの定性的および定量的組成の形成は、生物的および非生物的環境要因の影響下で発生します。
生産サイクル中の群集の構造的および機能的指標の変化は、1970 年のカンダラクシャ湾の生物学的季節の分析の例によって追跡できます (Fedorov および Bobrov、1977、Bobrov、1982)。
1970 年 4 月 15 日から 10 月 26 日までの観測期間中に、合計 51 個の観測点が設置されました(週 2 回)。 生成物は、水平線 0.2.5、5、10、25、50、75 m および底部のフラスコの放射性炭素法を使用して測定されました。 総日射量は、水中改造したジャニシェフスキー日射計を使用して測定されました。 対応する地層からの一次生産量を決定する実験と同時に、植物プランクトンの測深定量収集といくつかの水文学的および水化学的パラメーターの研究が実施されました。 光合成プロセスの効率を計算する際、これらの一次生産量の測定値は、次の計算に基づいてエネルギー単位で表されました。1 モルの CO2 を固定するのに 112 カロリーの光エネルギーが消費され、この値は 9.36 cal/mgC に相当します。
植物プランクトンの個体数から重量特性への移行は、次の式に従って行われました。
この式は、乾燥藻類バイオマスには 35% の灰分が含まれるという事実を考慮して得られました (Fedorov et al., 1974)。
1970 年の生育期の 1 日あたりの放射線量の値は、20.6 ~ 2300 cal/(cm" day) の範囲でした。5 月の最初の 10 日間で BOAy に浸透する日射量の増加は 100.6 ~ 181.4 cal/( cm" 日)は、氷の覆いが崩れるのと一致します。 総太陽エネルギーの値のわずかな減少は、気象条件によって引き起こされました. 気象条件の改善と日照時間の長さの増加により、光の最大値 (2300 cal/(cm. day)) が実際に一致するようになりました。その後の 1 日当たりの太陽放射量の低下はかなり滑らかな性質を持ち、LEHI HEM 持続時間 CBPTOBOTO IH H H BECOTE CONHL kl の減少によって引き起こされ、成長期の終わりまでに太陽エネルギーの量が減少します。 20cal/(cm.day)まで下がります。
したがって、183 日間の観測期間全体のエネルギー値は合計 22.8 kcal/cm で表され、秋の「氷下春」の時期の未知の透過日射エネルギーを外挿して求め、算出した。 22kcal/cmまで。」 シーズン中、合計で 25 kcal/cm が貯水池に浸透しました。」 冬の間、氷の下に浸透する太陽放射エネルギーの量はわずかであるため、後者の値は年間量と考えることができます。分布の研究成長期の水柱内の太陽エネルギーの計算により、水による光の吸収係数 k=0.37-002 を計算することができました。
成長期の地表の温度は-1.54~15.65℃の範囲でした。 温度変動は深さとともに平準化されます。 氷が壊れた後の春の期間は、均質性の違反と明確に定義された層別の確立によって特徴付けられます。 温度ジャンプゾーンは、浸透する太陽エネルギーの増加とそれに対応するVMの加熱を伴い、表面からより深い層へと徐々に移動しました。 太陽エネルギーが最大の期間中、水温躍層は深さ 25 m に位置し、9 月末までこの地平線に留まりました。 10月には、気温の上昇がより深くなり、恒温傾向が明らかになりました。
テーブル内 表 5.1 は、季節的な LHHaMHK.H の分析に基づいて得られた、白海のいくつかの非生物的特徴を示しています。
プランクトンの栄養発達とその生産の分析HEIM Xapa KTepH cTH Kにより、V.G. Bogorov (1938)によって生物学的な春、夏、秋と呼ばれる植生の主要な期間を特定し確立することが可能になります。 白海は 3 つの時期すべてに特徴があります。 1970 年のカンダラクシャ湾滞在期間はそれぞれ 48 日、92 日、43 日でした。 
生育期のさまざまな期間における生産プロセスの絶対値を特徴付ける結果を表に示します。 5.2. P/Sent 係数は、Bice TO CTO,JI (5a B OIE bil. 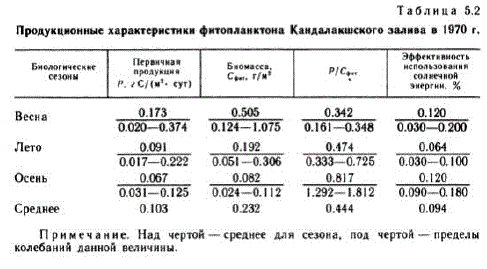
注記。 線の上は季節の平均、線の下は特定の値の変動範囲です。
生物学的な春の始まりは 4 月の最初の 10 日間に記録され、春の氷河下クライマックス (Fedorov、1970a; Kokin、Koltsova、1972) は約 2 週間続き、安定した P/Cphi 係数によって特徴付けられます。 藻類のエコニッチの分布は、第 2 のマッカーサー モデル (Kokin and Koltsova、1972) に対応します。
春のバイオマスと藻類の豊富な発生は氷の下で始まり、氷が壊れた後に最大に達します。 植物プランクトンは、BH La MH JULHETOMOBOTO KOM TIJEKCE: Fragilaria Oceanica、Chaetoceros holsa ficus、Nitzschia frigida、Maticula septentrionalia によって代表されます。 最も豊富な種は F. oceapisa でした。 光層は栄養素が豊富で、植物プランクトンの総バイオマスの 86% を含んでおり、光合成速度と同様に、この期間中に生育期全体の最大値に達します (表 5.2 を参照)。 動物プランクトンによる藻類の消費(Konoplya、1973)は、藻類バイオマスの減少と栄養素をめぐる食物競争の減少につながりますが、藻類の分布はすでに最初のマッカーサーモデルに対応しています。
珪藻の数の増加によって引き起こされる植物プランクトンの発生の夏の最大値(最初)は、その数が減少した後、ペリジニウム藻類によって「拾われ」、2番目の夏の最大値を形成します。 7月中旬から8月末にかけて、安定したP/Cフィット率で夏のクライマックスが確立されます。 生物学的な夏の主な形態は、SceletopemacostalumおよびChaetoceros属の種(主にCh.compostis)です。 この期間の一次生産量は、平均値 0.091 g C/(m"、日)、バイオマス - 0.192 g C/m" によって特徴付けられます。 栄養素をめぐる栄養競争が展開され、
秋の植物プランクトンの発生は弱いです。 8月下旬から9月中旬にかけてバイオマスee、Rb Ae/y!"b、pyAhQの成長が観察されます。 9 月 20 日の一次生産量が 0.125 g C/(m"、日) および P/Seite 係数 (最大 1812) に増加しただけで、植物プランクトンの短期的な活性化が示されています。この期間中、Dinophyes powegica の発生が見られました。生物学的秋の寒さを好む優勢植物が観察されます (Fedorov, 1969). 秋の遷移期間は生態系を短期的な最高潮に導きます, それは 9 月末まで続きます. 気温と日射量のさらなる低下,同様に、密度ジャンプの中断による混合層の増加は、植物プランクトンバイオマスの深さの再分布と生成物に対する破壊プロセスの優位性をもたらします。
一般に、このような植物プランクトン生産サイクルの経過は、海の他の地域でも予想されます。 考えられる違いは、植物プランクトンの生育期の始まりと終わりのタイミング、および主要な藻類種の定性的組成におけるいくつかの違いに表れ(Zhitina、1981)、間違いなく植物プランクトン生産プロセスの結果に影響を与えるでしょう。水域のこれらの部分にあります。
1970 年の植物プランクトンの生産サイクルの研究から得られたデータにより、白海の栄養性の分類を明確にすることが可能になりました。 追加の特性として、植物プランクトンによる太陽エネルギーの利用効率が使用されました (表 5.2 を参照)。これは植物プランクトンの状態の不可欠な特性であり、一連の環境条件に依存し、植物プランクトンのバイオマスとの直接的な関係は示されていません。 (フェドロフ、ボブロフ、1977)。
植物プランクトンの主な生産特性の計算値を、生産性の異なる海域の既知のデータと比較すると(Sorokin、1973)、カンダラクシャ湾は植物プランクトン生産レベルが0.05〜0.10 gの非生産地域に属していると結論付けることができます。 C/(m* - day) および太陽エネルギー利用効率は 0.04% であり、貧栄養水域の特徴です。 したがって、カンダラクシャ湾は、O.I. Koblenz-Mishke の分類 (Koblenz-Mishke et al., 1970) によれば、深層水の混合がほとんどなく、栄養クラス 1 として分類できます。
表 6.7 ~ 6.9 は、さまざまなサンゴ礁の水域における一次生産量の測定結果をまとめたものです。 表からわかるように、同じサンゴ礁の海域内でも一次生産量は大きく異なります。 しかし、これらの後者の変動は極めて自然であり、主にサンゴ礁の浅いゾーンからラグーンの中心に向かう部分における植物プランクトンの濃度の変化に関連しています。
一次生産量が少ない環礁(ラウ島およびマーシャル諸島、ツアモツ諸島の環礁)のラグーンでは、下限値(1 日あたり 2 ~ 4 mg C/m 3)は貧栄養条件下での値に近いです。熱帯海域の貿易風がそれらを洗い流しており、通常は 1 日あたり 1 ~ 5 mg C/m 3 です (Sorokin, 1971a)。 しかし、そのような環礁であっても、ラグーンの水域では植物プランクトンの発生が季節的に発生する可能性があり、それは「ブルーム」の強さに達する可能性があります(Sournia、Richard、1976a)。 表のデータからわかるように。 6.7、環礁ラグーンでは、植物プランクトンの発生レベルが極めて低い貧栄養熱帯海域に洗われているラグーンであっても、一次生産量はほとんどの場合、中栄養海洋盆地における植物プランクトン生産性の平均レベル、場合によっては上限レベルに相当します(15~50 mg C/m 3/m 3 )。日)。 インド洋の南貿易風流の非常に非生産的な海域の中心に位置するカルガドス・カラホスの大きな環状礁のラグーンでは、藻類バイオマス(主に珪藻)による一次生産量が1日あたり250 mg C/m 3 に達しました。 )5.2 g/m 3 を超える(表 6.10)。 ファニング環礁 (ラウ諸島) の閉鎖礁湖では、渦鞭毛藻と球状藍藻の発達期に、高い生産量 (1 日あたり 70 ~ 100 mg C/m3) が記録されました (Gordon et al., 1971)。 この環礁は貿易風流の貧栄養海域にも位置しています。 したがって、貧栄養水の流れの中に位置する個々の環状礁のラグーン内でも、非常に生産性の高い植物プランクトン群集が存在する可能性があり、その生産量とバイオマスは、それらを洗浄する海水よりも1〜2桁高くなります。
利用可能ないくつかの測定結果から判断すると、外洋の環状礁のラグーン水中の光合成活性のあるクロロフィル a の濃度は、周囲の海水より平均して 5 ~ 10 倍高いです。 単一の測定によると、異なるサンゴ礁のラグーンでは 0.2 ~ 1 mg/m3 の範囲で変動しました。 タカポト環礁のラグーンでは、長期観察の結果 (Sougnia、Ricard、1976a) によれば、クロロフィル濃度は 0.26 から 0.77 mg/m 3 の範囲で変動し、ラグーンの異なる部分では環境に応じて顕著に変化しました。満潮時の海水への侵入の強さ。
植物プランクトンとその産物の研究に多大な貢献をしたのは、大きな島々の近くの堰堤や海岸のサンゴ礁の水域で行われた季節観察でした。そこでは、サンゴ礁と隣接する海洋のネライトゾーンの生態を研究するために生物学的ステーションが設置されました。 このようなよく研究されたサンゴ礁の中で、ノシベ島沖およびトゥレア沖(マダガスカル)のサンゴ礁、オアフ島沖のハワイアンリーフ、オーストラリアのグレートバリアリーフ、ワプラオ、ティアウラ、およびサンゴ礁について言及する必要があります。ニャチャン近くのベトナム中部のサンゴ礁、タヒチ島とモーレア島にあるファアア。 堰堤と海岸礁の一次生産量と植物プランクトンバイオマスの測定結果を表にまとめます。 6.8、6.9、6.10: そこに示されているデータは、調査対象のほぼすべてのサンゴ礁で大きく異なります。 これらは、同じサンゴ礁の異なるゾーンにおける一次生産量の定期的な測定値を反映しています。 同時に、その最小値(および植物プランクトンバイオマスの最小値:図6.10)は、水が底層個体群および藻類叢と集中的に接触しているサンゴ礁の浅水ゾーンに対応します。サンゴ礁の。 これらのゾーン(平地、パッチ礁、沿岸)では、通常、植物プランクトンが枯渇し、その生産量がここで急激に減少します。 これらの表に示されている各サンゴ礁の植物プランクトン生産量とバイオマスの最大値は、水が入っていない深さ 15 ~ 20 m を超えるラグーンまたは周縁礁または沿岸礁の外側斜面のゾーンに対応しています。サンゴ礁の底の人々と直接接触します。 これらのゾーンの一次生産量は、原則として、隣接する外海の水域よりも大幅に高くなります。 平均して、環礁のラグーンや外洋の環状礁の水域よりも高いです。
マダガスカル沖のサンゴ礁、セイシェル諸島、ソコトラ島など、インド洋の多くの海岸および堰礁の水域で、富栄養水域のレベルに達する高い一次生産量が確認された。アオコの「開花」期に、B.バリアリーフのラグーンで観察されました(図6.9を参照)。 ここでの毎日の生産量は、水の上層で 1 日あたり 50 ~ 300 mg C/m 3、または 0.7 ~ 2.8 g C/m 3 に達しました。 さらに、貧栄養の海水によって洗われた島々(タンギ、オアフ島、トカゲ)のバリアリーフの水域では、ラグーンでの一次生産量は 50 ~ 60 mg C/m 2、または 1 m 2 日あたり 0.4 ~ 0.7 g の炭素に達することがあります。 、これは中栄養水の生産性の上限に相当します。 ベトナム中部の海岸と南シナ海の島々(トゥー諸島)およびタイ湾(ナムズー諸島)の岩礁地帯(表 6.8、図 6.10 を参照)では、主要な植物プランクトンの値が観察が植物プランクトンの季節的最小期間中に行われたという事実にもかかわらず、生産量は高かった(Sorokin et al., 1982)。 一次生産量の最高値は、原則として、サンゴ礁から10〜100 m離れた深さ5〜15 m以上のサンゴ礁の外縁の水域で観察されました(図6.10を参照)。 通常、1 日あたり 30 ~ 40 mg C/m 3 を超えました。 1984 年のこのゾーンの個々のセクションでは、1 日あたり 100 mg C/m 3 を超える一次生産量が記録されました。
海岸および堰礁における植物プランクトンの生物量に関する情報は依然として非常に断片的です。 シーズン中の植物プランクトンのバイオマスの最も詳細な測定は、ノシベ島 (マダガスカル) の Angot (1968) によって行われました。 これらの測定結果を図にまとめます。 6.9. テーブル内 表 6.10 は、インド洋と太平洋の多くの大きなサンゴ礁における植物プランクトンのバイオマスに関する文献で入手可能なデータを示しています。 表からわかるように、堰堤や海岸のサンゴ礁の水域における植物プランクトンのバイオマスは 3 ~ 8 g/m3 に達することがあります。 最も低い値は、沈下状態にあった太平洋中央部の沿岸および堰礁(ニューヘブリディーズ諸島のパナロラ島およびパアマ島沖の岩礁、ラウ諸島のツヴタ島沖の岩礁)の水域で観測されました。 )。
堰堤および沿岸礁の水域におけるクロロフィル a の含有量に関する情報も同様に限られています (表 6.11)。 最も詳細な測定は、フランスの研究者によってモーレア島とタヒチ島のラグーンの水域で行われました (Ricard、1976; Sournia、Ricard、1976)。 これらの測定により、一年のさまざまな季節におけるクロロフィル濃度は 0.03 ~ 0.17 mg/m 3 の範囲内で変化することが示されました。 オアフ島沖のハワイアン リーフのラグーンと、冬の B. バリア リーフのラグーンでは、クロロフィル濃度は中栄養海水のそれに近い 0.1 ~ 0.3 mg/m3 でした。 秋から夏にかけて、B.バリアリーフのラグーンおよびノシベ島(マダガスカル)沖のサンゴ礁の水域で植物プランクトンが最も活発に発達した時期には、水中のクロロフィル濃度は以前よりも高かった。 1 mg/m 3 で、これは富栄養水の含有量に近い値です (Angot、1968)。 防波堤ゾーンにおける微生物底生生物が豊富な底質の全層の再懸濁がサンゴ礁の水中のクロロフィル濃度に及ぼす影響は確立されている(Walker、1981)。
上で述べたように、植物プランクトンの一次生産量と現存量の値は、同じサンゴ礁のゾーンによって大きく異なります。 ほとんどの場合、バイオマスと植物プランクトンの生産量の最低値は、サンゴ礁の浅いゾーン、特に満潮と干潮時にサンゴ礁を通過する水が海底生物と最も接触するサンゴ礁平坦部の特徴です。サンゴ礁。 平地の浅いエリアでの植物プランクトンの減少の理由は、サンゴ礁の底層フィルターフィーダーによる植物プランクトンの消費に見られるはずです (Glynn、1973a)。 さらに、Nitzschia delicatissima や Chaetoceros の分枝細胞などの動物の細胞は、サンゴ礁の乱流水中で、特にサンゴ礁の硬い表面や浮遊粒子と衝突したときに損傷する可能性があります。 ラグーンのより深い部分やサンゴ礁の外縁を越えた水域では、水とサンゴ礁との接触が減少し、サンゴ礁の底部群集への一部の栄養素の放出により植物プランクトンの発生が激化します。
バイオマスと生産量の定量的指標、およびクロロフィル濃度の観点から見ると、サンゴ礁地帯の植物プランクトンは外洋の周囲の海域よりも豊富です (Michel, 1969; Sorokin, 1971; Ricard, 1976; Sournia, Ricard 、1976a)。 図 6.3 は、ヘロン リーフ (B. バリア リーフ) から外洋までのセクションに沿った植物プランクトンの一次生産量の変化の典型的な図を示しています。 すでに6〜9 kmの距離では、それは1桁以上減少します。
サンゴ礁の深い部分にある白い円盤に沿った水の透明度は通常非常に高く、平均で 10 ~ 15 m ですが、場合によってはそれより低い場合 (カントン環礁では 5 メートル) や、30 メートルを超える場合もあります (マジュロ環礁) )。 したがって、植物プランクトンの光合成は、実際にはその厚さ全体にわたって光によって制限されません。 図では、 図 6.11 は、平均透明度 (10 ~ 18) のインド洋のいくつかのサンゴ礁の海域における入射放射線の典型的な減衰曲線を示しています。 この図からわかるように、植物プランクトンはラグーンの最底部でのみ光不足を経験する可能性があります。
植物プランクトンの季節変動の例を図に示します。 6.9、6.12、6.13。 彼らは、一年のさまざまな季節にその組成が変化するだけでなく(6.2.1を参照)、そのバイオマスと機能的活動にも変化があることを示した。 季節中のサンゴ礁における一次植物プランクトン生産量とクロロフィル濃度の変動は桁違いに達した(Angot、1968; Sournia、1972)。 さらに、貧栄養水域に位置するタカポト環礁のラグーンでも、季節最大時のクロロフィル濃度は中栄養水域に特徴的なレベルに達した(Sournia and Ricard, 1976)。 堰堤と海岸の礁域の水域では、その濃度は富栄養海域の含有量に相当する 1 mg/m3 近くであり、ノシベ島沖の岩礁帯の植物プランクトンの現存量は 3 g/m3 を超えていました。 タウンズビル近くのオーストラリアンバリアリーフの B. ラグーンでは、水中のクロロフィル濃度の顕著な季節変動 (0.13 から 0.5 ~ 2.5 mg/m 3 まで) が観察されました (Sammarco、Crenshaw、1984)。 最大値は12月から4月までの夏から秋にかけて観察されました。 それは、雨が降って排水面が増加した時期と一致しました(図6.14)。 この期間中、ラグーンの水域では植物プランクトンの集中的な発生が観察され、そのかなりの部分が藍藻類のトリコドクスミウムでした (Relevante et al., 1982)。 植物プランクトンの発達に対する川の流れの刺激効果は、ニューカレドニア沖のサンゴ礁水域でも注目されました (Binet、1984)。 提示されたデータは、温帯海域とは異なり、熱帯海域のプランクトン群集はある種の恒常性の状態にあり、重大な季節変化を経験しないという理論(Parsons et al.、1977)の根拠がないことを示しています。 このような考えは、明らかに熱帯海域における適切な季節観測サイクルの欠如の結果でした (Sournia、1969)。 熱帯沿岸海域で植物プランクトンの発生が季節的に最大になる理由は、主に、サンゴ礁に隣接する海域の水循環の変化によるものであり、これには年周期性があります。 さらに、これらの変化は、熱帯地域であっても、ほとんどの場合、秋と春に起こります。 植物プランクトンの発生の季節的最大値が秋または春に記録されるのは偶然ではありません。
サンゴ礁海域における植物プランクトンの発生を研究する過程で、その群集の構成と存在量、およびその生産力に大きな日次変動が存在することが証明された(ミリマンとマンケン、1972年; ソーニアとリカール、1976年; ソロキン) 、1979)。 このような変動は外洋の植物プランクトン群集でも発生します。 そこでは、植物プランクトンの成長が毎日周期的に行われることと、下層の水層から受光層に移動する動物プランクトンによる夜間の藻類の消費が激化することが原因で発生します。 同じ理由がサンゴ礁地帯の沿岸海域でも作用し、植物プランクトンの発生に毎日の周期性を引き起こしています。 しかし、これらの海域では毎日の潮の干満の影響も受けます。 南豆諸島付近のサンゴ礁での毎日の観察結果が示すように、サンゴ礁の動物プランクトンが夜間に水柱に移動する植物シャンクトンの放牧の影響で、早朝の一日の生産量は一桁減少します。 。 しかし、午前11時までに、特に植物プランクトンが集中的に繁殖するサンゴ礁の端では、植物プランクトンの個体数は大幅に回復します(図6.15)。
外洋の周囲の水域と比較して、サンゴ礁海域でのバイオマスと植物タンクトンの生産が増加している理由は何ですか? この問題は、特に、このような増加が外洋とあまり変わらない非常に低い栄養素の絶対濃度で発生するという事実や、フィルターによる植物プランクトン群集の異常に集中的な利用の条件下で発生するという事実を考慮すると、非常に複雑です。 -サンゴ礁の動物相に餌を与える。 この問題に関して表明された意見は非常に矛盾しています (Sournia, Ricard, 1976; Sournia, 1976b; Salvat 1982)。 ほとんどの著者は、いわゆる「島嶼質量効果」を主な理由として指摘しています。 この効果は、島の質量と入ってくる海流の流れとの相互作用により、島の沿岸域の水が栄養分で富み、その結果、深層水が表面に上昇することにあります。 例えばマーキス諸島など、貿易風流の中心に位置する島々におけるネライトゾーンの濃縮の影響(Sournia、1976c)。 B. バリアリーフ (Gilmartin、Relevante、1974)、ハワイ (Doty、Capurro、1959)、セイシェル (私たちの観察) は間違いなく発生します。 その影響下で、そのような島のネライトゾーンでの一次生産は非常に高く、それがサンゴ礁水自体の生産性のレベルに影響を与えます。 しかし、こうした条件下であっても、サンゴ礁ラグーンの水域における植物プランクトンの生産量とバイオマスは、通常、サンゴ礁ラグーンに隣接し、それらとの集中的な交流が行われているネライトゾーンの水域よりも高い(Marshall, 1965; Michel, 1969)。 ;Sournia、Ricard、1976、1976a;Thompson、Golding、1982)。 これらのデータを考慮すると、サンゴ礁海域における植物プランクトンの生産性が増加する主な理由は、底質および浮遊従属栄養生物によって生物起源要素(主に窒素、リン)が水柱に継続的に放出されるためであると結論付けることができます。サンゴ礁で大きな力で起こる有機物の破壊プロセスの結果です。 同時に、サンゴ礁の水柱における栄養塩の代謝回転速度が非常に高いため、隣接する海と比較してサンゴ礁地帯の水域における塩分栄養素の絶対濃度の増加を常に記録できるわけではありません。水域(図5.1)。
植物プランクトンに対する光と温度の影響。
コンスタンチノフの研究によると、水温と貯水池の表面に到達する太陽放射が、藻類のエネルギーと新しい有機物 (OM) を形成する能力を決定する最も重要な要素です。 独立栄養生物の代表である藻類にとって、光は最も重要な要素です。 それは光合成、成長、発達を決定します。 変化する光条件に適応する過程で、植物の光合成装置は放射エネルギーを最大限に活用するように調整されます。
コンスタンチノフは、太陽放射の吸収率と水への光の浸透の度合いは、地理的緯度、季節、時刻によって変化する太陽の高さ、水に溶けている有機物質の量に依存することを証明しました。水、水の色、濁り、貯水池の表面の状態。 風がない場合、入射放射線の 5% が水面で反射され、風の強い場合は 15%、風が強い場合は 30% になります。 光の強さは深さとともに減少します。 透明度 1 ~ 2 m の湖や貯水池では、入射放射線のエネルギーの 5 ~ 10% が 1 m、2 m より深い深さまで浸透します (0.015 ~ 0.04 J/cm 2 - 分)。
M.R. グセフは、ラン藻は他の藻類グループに比べて光をあまり要求しないと考えています。 光は、藻類の光合成、成長、発達を決定します。 さまざまな植物プランクトン生物の光要件は種ごとに異なります。 緑藻および藍藻は光を必要とすると考えられています。 最大限の光合成を実行するには、珪藻や緑藻に比べて 1.2 ~ 2 倍少ない日射強度が必要です。 クズネツォフ氏は、珪藻は照明に対する要求が少なく、明るい表層を避け、透明度の低い貯水池では深さ2~3メートル、透明な海水では深さ15~20メートルに生息していると指摘する。 緑藻類17では、光飽和は5〜7千ルクス、珪藻では10〜2万ルクス、渦鞭毛藻では25〜3万ルクスで発生することが確立されています。 これは、珪藻が必要とする光の量が少ないという事実と矛盾します。 湖での珪藻のブルームは、細胞が水柱全体を循環する春や秋の混合サイクル中など、光条件が非常に変化し、放射線レベルが低いときによく発生します。 V.N. Goponenko が指摘しているように、単細胞藻類では 6 ~ 8,000 ルクスで光飽和が発生します。 1,000ルクスの照度で生育した藻類培養物では、緑藻類では5〜7.5千ルクス、珪藻では1〜2,000ルクス、ペリディニウムでは25〜3万ルクスで光飽和が発生します。 によると
N.P. Kalinichenko、珪藻 Stephanodis cushantzschii の最適照度は、日照時間 12 時間と 16 時間で 2.6 千ルクス、Asterionella formosa の場合、同じ露出で 5 千ルクスです。
貯水池の観察期間中、上部ポクロフスコエ湖の水温とpHは20日ごとの間隔で測定されました。 得られたデータを表 6 に示します。また、どの季節にそれらが最大量に達するかを示すために、いくつかの代表的な緑藻および藍藻についてグラフも分析および編集しました。
表 6 - 2013 年から 2014 年の温度条件と pH インジケーター。
図 3 から、緑藻 (Ankistrodesmus acicularis) が夏に優勢で 51% を占め、冬にはその量が 2% であることがわかります。 このような指標は、夏の平均気温が26.6度であり、発達に有益な効果があるという事実によるものです。
図 4 では、別の代表的な緑藻 (Scenedesmusquadricauda) が春に優勢で 42% を占めますが、冬にはその量が 3% を超えず、一年を通じてすべての季節で見つかりました。
図 3 - 2013 年のポクロフスコエ湖の緑藻類 (Ankistrodesmus acicularis) の数 (年間総価値の%)

図 4 - 2013 年のポクロフスコエ湖の緑藻類 (Scenedesmusquadricauda) の豊富さ (年間総価値の %)
図4は、夏に優勢な藍藻(Microcystis aerugenosa)の量が70%を占め、夏の平均水温は26.6度と、その生育に好条件であることを示しています。 冬にはそれほど多くはなく、約1%です。
図 5 は、藍藻 (Oscillatoria tenui) が夏に優勢で 63% を占め、冬には見られないことを示しています。

図 5 - 2013 年のポクロフスコエ湖における藍藻類 (Microcystis aerugenosa) の豊富さ (年間総価値の %)

図 6 - 2013 年のポクロフスコエ湖の藍藻類 (Oscillatoria tenuis) の豊富さ (年間総価値の %)
図 7 は、珪藻 (Navicula platystoma) を示しています。この珪藻は、秋の平均水温が 17.5 ℃である秋の期間に 48% の最大存在量に達し、春の平均水温が 13 ℃である秋の期間に 35% に達します。 。

図 7 - 2013 年のポクロフスコエ湖の珪藻 (Navicula platystoma) の数 (年間総値に占める割合)